The Ganoidei
In former periods of the world's history, this was one of the largest and most important of the orders of fishes; but, at present it comprises only the seven genera - Lepidosteus, Polypterus, Calamoichthys, Amia, Accipenser, Scapirhynchus, and Spatularia, which are either partially or wholly confined to fresh water, and are found only in the northern hemisphere. These fishes differ very widely from one another in many points of their organization, but agree in the following characters, some of which they possess in common with the Elasmobranchii and others with the Teleostei, Thus:- The bulbus aortae is rhythmically contractile, and provided with several rows of valves, as in the Elasmobranchii.
- The optic nerves unite in a chiasma, as in the Elasmobranchii.
- There is a well-developed spiral valve in the intestine, as in the Elasmobranchii, in all but Lepidosteus, which possesses only a rudiment of such a valve.
On the other hand:
- The branchial processes are not fixed throughout their extent to the wall of a branchial sac, which extends beyond them, as in the Elasmobranchii; but their extremities project freely beyond the edge of the septum which separates each pair of branchial clefts, as in the Teleostei; and, as in the Teleostei, they are covered by a bony operculum.
- There is a large air - bladder connected by a permanently open pneumatic duct with the oesophagus, as in many Teleostei.
- As in the Teleostei, there is no cloaca.
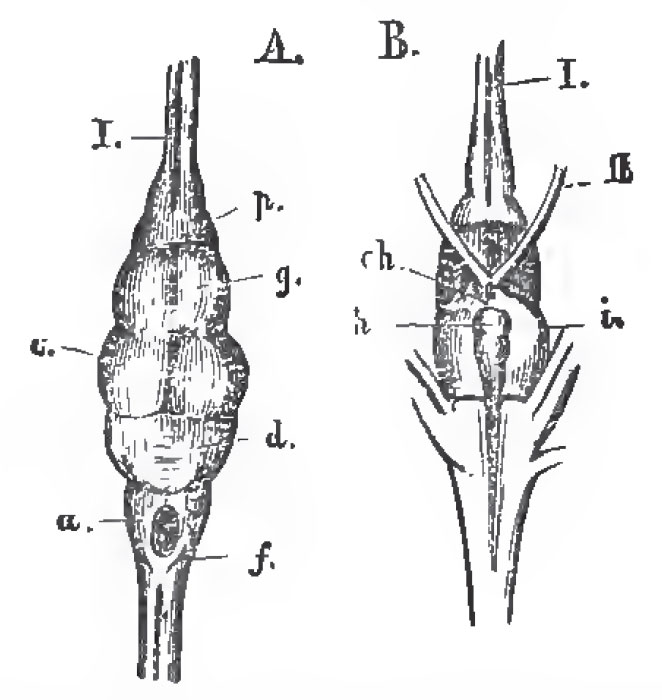 |
| Fig. 38. - The brain of Lepidosteus semiradiatus. A. From above; B. From below. f the medulla oblongata; d, the cerebellum; c, the optic lobes of the mesencephalon. o, the cerebral hemispheres; h, the pituitary body; i, the lobi inferiores. Ch, the chladma; I., olfactory; II., optic nerves. |
The exoskeleton presents the most extreme variations in the Ganoidei. Spatularia is naked; Accipenser and Scapirhynchus develop numerous dermal plates composed of true bone; Amia is covered with overlapping cycloid scales; Lepidosteus and Polypterus have solid, rhomboidal, enamelled scales, which not only overlap, but are fitted together by pegs and sockets, where their anterior and posterior edges come into contact.
The endoskeleton is not less diversely modified; and it is worthy of remark that no sort of relation, either direct or inverse, is traceable between the completeness of the endoskeleton and that of the exoskeleton. Thus Spatularia, Scapirhynchus, and Accipenser have a persistent notochord, in the sheath of which mere cartilaginous rudiments of the arches of vertebrae appear. The ribs, when present, are partially ossified. Polypterus and Amia have fully ossified vertebrae, the centra of which are amphicoelous. Lepidosteus also has fully-ossified vertebrae; but their centra are opisthocoelous, having a convexity in front and a concavity behind, as in some Amphibia.
More or fewer of the anterior vertebrae, or their cartilaginous representatives, are united with one another, and with the posterior part of the skull. And the cranium may consist principally of cartilage, membrane bones being superadded; or the primordial cartilage maybe largely superseded by bone, as in the Teleostei.
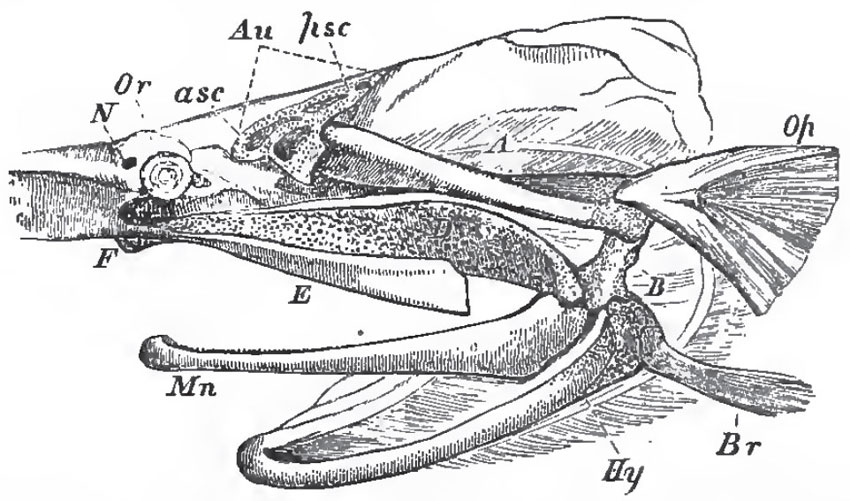 |
| Fig. 39. - Side-view of the skull of Spatularia, with the beak cut away, and the anterior (asc), and posterior (psc), semicircular canals exposed: Au, auditory chamber; Or, the orbit with the eye; N, the nasal sac; Hy, the hyoidean apparatus; Br, the representatives of the branchiostegal rays; Op, operculum; Mn, mandible; A, B, suspensorium; D, palato-quadrate cartilage; E, maxilla. |
The framework of the jaws in Spatularia is very similar to that in the Elasmobranchii. There is a partly cartilaginous, and partly ossified, suspensorial cartilage (A, B, Fig. 39), which gives attachment below, directly, to the hyoidean arch (Hy), and, indirectly, to the jaws. The latter consist of a palato-quadrate cartilage (D) united by ligament with its fellow, and with the prefrontal region of the skull at F.; and presenting, at its posterior end, a convex articular head to the cartilage of the mandible, or Meckelian cartilage, Mn. It is obvious that A, B, corresponds with the hyomandibular, or suspensorial, cartilage in the Sharks and Rays; D, with the palato-quadrate cartilage, or so-called "upper jaw," and the cartilage of the mandible with the lower jaw in these animals. But, in the Ganoid fish, an osseous operculum (Op) is attached to the hyomandibular; and a branchiostegal ray (Br) to the more strictly hyoidean part of the skeleton of the second visceral arch; while a membrane bone (E) representing the maxilla, and another (Mn) the dentary, of the lower jaw is Teleostei, are developed in connection with the palato-quadrate and mandibular cartilages.
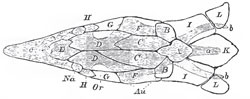 |
| Fig. 40. - The cartilaginous skull of a Sturgeon, with the cranial bones. The former is shaded, and supposed to be seen through the latter, which are left unshade. l: a. ridge formed by the spinous processes of the anterior vertebrae; b, b, lateral wioglike processes; c, rostrum; Au, position of the auditory organ; Na, position of the nasal sacs; Or, that of orbit. The membrane bones of the upper surface are: A, the analogue of the supra-occipital; B, B, of the epiotics; E, of the ethmoid; G, G, of the postfrontals; II, II, of the prefrontals; C, C, the parietals; D, D, are the frontals, and F, F, the squamosals; K, the anterior dermal scute; I, I, and L, L, dermal ossifications connecting the pectoral arch with the skull. |
In Lepidosteus, Polypterus, and Amia, the skull presents not only membrane bones, but, in addition, basi-occipital, ex-occipital, and prootic ossifications of the primordial cartilage, to which others may be added. The vomers are double, as in the Amphibia (? Polypterus). The apparatus of the jaws has become modified in accordance with the Teleostean type of structure. The suspensorium consists of two ossifications united by a cartilaginous intermediate portion. The upper- broad, and movably articulated with the periotic capsule-is the hyomandibular; the lower answers to the symplectic of osseous fishes. The cartilaginous palato-quadrate arcade is, in part, replaced by a series of bones: the palatine lies in front, and is connected with the prefrontal region of the skull; behind it, lie representatives of the pterygoid, the metapterygoid, the ectopterygoid; and, most posteriorly, of the quadrate bone. The last furnishes a condyle to the articular element of the mandible. The symplectic is either loosely connected with the quadrate, as in Lepidosteus. or more closely united with it, as in the other genera.
In Lepidosteus and Amia, a strcng and long membrane bone, the preoperculum, is developed on the outer side of the hyomandibular and quadrate bones, and connects them still more firmly together.
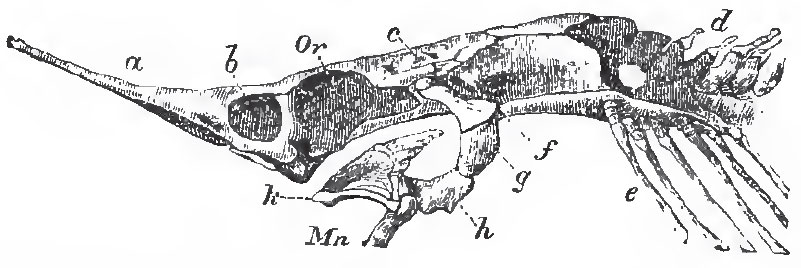 |
| Fig. 41. - Side-view of the cartilaginous cranium of Accipenser: a, rostrum; b, nasal chamber; Or, orbit; c, auditory region; d, coalesced anterior vertebrae; e, ribs; f, g, h, suspensorium; k, palato-maxillary apparatus; Mn, mandible. |
The maxilla is represented by a series of small separate ossifications in Lepidosteus. The proximal end of the mandibular cartilage ossifies, and becomes a distinct articidare. A dentary element is added on the outer, and a splenial one upon the inner side of the cartilage; and in Lepidosteus, angulur, supra-angular, and coronary elements are added, so that the components of the mandible are as numerous as in reptiles. Lepidosteus and Amia have branchiostegal rays, but Polypterus has none - at any rate, of the ordinary kind. A single jugular plate is developed between the rami of the mandible in Amia, and there are two such plates in Polypterus, which may possibly represent branchiostegal rays.
In Accipenser, Spatularia, and Amia, the pectoral arch presents two constituents: one, internal and cartilaginous, answers to the cartilaginous pectoral arch of the Elasmobranchii, and to the scapula and coracoid of the higher Vertebrata; the other, external, consists of membrane bones representing the clavicular, supra-clavicular, and post-clavicular bones of the Teleostei. In Lepidosteus one centre of ossification appears in the cartilage; in Polypterus, two. The upper represents the scapula, and the lower the coracoid.
It has been already stated (p. 38) that Polypterus comes nearest to the Elasmobranchii in the structure of the rest of the limb. The numerous dermal fin-rays, all nearly equal in size, are connected with the rounded periphery of the broad and elongated disk formed by the skeleton of the fin; and the scaly integument is continued to the bases of the fin-rays, which thus seem to fringe a lobe of the integument. Hence the fin is said to be lobate. In the other genera, only two of the basal cartilages are present, and some of the radialia come into contact with the shoulder-girdle between them. In addition, the anterior dermal fin-ray is much larger than the others, and becomes directly connected with the anterior basal cartilage. Thus, in the structure of their fins, as in so many other characters, the Ganoidei are intermediate between the Elasmobranchii and the Teleostei.
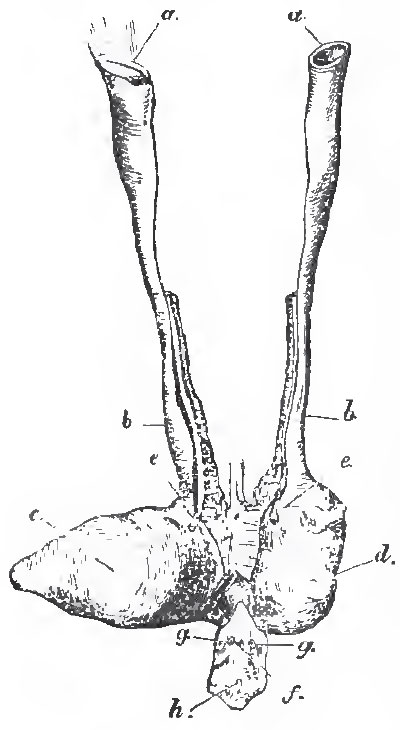 |
| Fig. 43. - The female reproductive organs of Amia Calve: a, a, the open ends of the genital ducts; b, b, oviducts; c, d, the right and left divisions of the urinary bladder, e, e, the openings of the ureters into the bladder; f, the anus; g, g, the abdominal porses; h, the urogenital sperture. |
In Accipenser and Polypterus, spiracula, or openings which communicate with the mouth, lie on the top of the head, in front of the suspensorium, as in many Elasmobranchs.
Lepidosteus, Accipenser, and Scapirhynchus, have branchiae attached to the hyoidean arch, as in the Elasmobranchii. They are now called opercular gills.
In Polypterus the air-bladder is double and sacculated, and the pneumatic duct opens upon the ventral aspect of the oesophagus. The airbladder thus becomes exceedingly like a lung; but its vessels are in communication with those of the adjacent parts of the body - not with the heart, as in a true lung.
In Lepidosteus, the ducts of the male and female reproductive organs are continuous with those bodies, and each duct opens into the dilated ureter of its side. In the other Ganoids the proximal ends of the genital ducts, in both sexes, open widely into the abdominal cavity. In Polypterus the united ureters open into the cavity of the confluent oviducts, while, in the other Ganiods the oviducts open into the dilated ureters. (Fig. 42.)
When the fossil, as well as the existing Ganoidei, are taken into account, they form a large order, divisible into the following sub-orders: 1 Amiadae, 2. Lepidosteidae, 3. Crossopterygidae, 4. Chondrosteidae, all of which have living representatives; while the other three - viz., 5. Cephalaspidea, 6. Placodermi, and 7. Acanthodidae - have been extinct since the Palaeozoic epoch, and are only ranged among the Ganoids provisionally, in as much as we have no knowledge of their internal anatomy.
1. The Amiadae have a single living representative in the rivers of North America - Amia calva; and it is not certain that any member of the group occurs in the fossil state. The cycloid scales, preoperoulum, single median jugular plate, branchiostegal rays, non-lobate paired fins, and heterocercal tail, diagnose the sub-order.
2. The Lepidosteidae have rhomboidal enamelled scales, a preoperculum, branchiostegal rays, non-lobate paired fins, and heterocercal tail. These are represented in the rivers of North America at the present day, and in tertiary formations, by Lepidosteus; in the Mesozoic rocks, by a great variety of genera - Lepidotus,CEchmodus, Dapedius, etc.; and, in the Palaeozoic epoch, by Paleoniscus in the Carboniferous, and probably by Cheirolepis, in the Devonian, formation.
3. In the Crossopterygidae the scales vary in thickness and ornamentation, and may be thin and cycloid, or thick and rhomboid. The dorsal fins are either two in number, or, if single, verv long, or multifid. The pectoral fins, and usually the ventrals, are lobate; they are sometimes rounded, as in Polypterus - sometimes greatly elongated and almost filiform, as in Holoptychius (Fig. 43). There are no branchiostegal rays, but two principal, and sometimes many smaller lateral, jugular plates. The tail may be either diphycercal or hetero cereal.
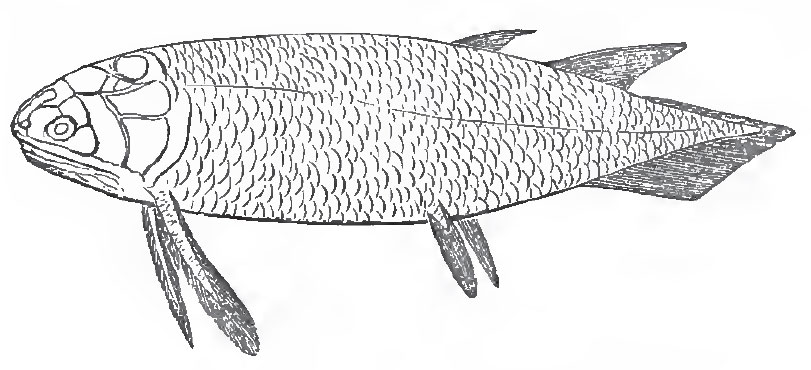 |
| Fig. 43. - Restoration of Holoptychius. |
The only living representatives of this sub-order are Polypterus and Calamoichthys, which inhabit the rivers of North Africa. Neither of these are known to occur in the fossil state. The only family of the sub-order at present known among Mesozoic fossils is that of the Caelacanthini, a remarkable group of fishes with a persistent notocbord, rudimentary ribs, an air-bladder with ossified walls, and a single interspinous bone for each of the two dorsal fins. The Caelacanthini also occur in the Carboniferous formation; and the great majority of the Crossopterygidae are found in this and the Devonian formations (Osteolepis, Diplopterus, Glyptolaemus, Megalichthys, Holoptychius, Rhizodus, Dipterus, Phaneropleuron, etc.). Megalichthys, Dipterus, and probably a few other of these fishes, have partially ossified vertebral centra; the rest possessed a persistent notochord. It is by the Crossopterygidae that the Ganoids are especially connected with the Dipnoi, and, through them, with the Amphibia.
4. The Chondrosteidae are either naked, or have dermal plates of bone in the place of scales. Neither the pectoral nor the ventral fins are lobate. The branchiostegal rays are few or absent, the tail is heterocercal. There are no cartilage-bones in the brain-case. The teeth are very small, or absent.
The Sturgeons (Accipenser) - which inhabit the northern rivers of Europe, Asia, and America, occasionally migrating to the sea - Spatularia, Aud Scapirhynchus (found in the rivers of North America), are the recent members of this group, which is represented, in the older Mesozoic rocks, by Chondrosteus.
5. The Cephalaspidae are remarkable fishes, probably allied to the Chondrosteidae, which occur only in the Lower Devonian and the Upper Silurian rocks, and are some of the oldest fish at present known. The head is covered by a continuous shield, which has the structure of true bone, in Cephalaspis, but more resembles certain piscine scales, in Pterapsis. The shield is prolonged into two horns at its posterolateral angles, and a median dorsal backward prolongation usually bears a spine, in Cephalaspis; the body is covered with flat bony scales or plates, and possesses two large pectoral fins. The characters of the body and fins of Pterapsis are unknown. Notwithstanding the excellent preservation of many of the specimens of these fishes, they have, as yet, yielded no evidence of jaws or teeth. Should jaws be absent, the Cephalaspidae would approach the Marsipobranchii more nearly than any of the other amphirhine fishes do.
6. The Placodermi, comprising the genera Coccosteus, Pterichthys, Asterolepis, and some others, are known to occur only in the Devonian and Carboniferous formations. In these fishes the pectoral region of the body is encased in great bony plates, which, like those of the skull, are ornamented with dots of enamel. The caudal region was covered with small scales in Pterichthys, while in Coccosteus it appears to have been naked. The pectoral member of Pterichthys is exceedingly long, covered with suturally-united bony plates, and united with the thoracic plates by a regular joint. In Coccosteus the pectoral member seems to have had the ordinary construction. The bones of the head and thorax of Coccosteus nearly resemble those of certain Siluroid fishes (e. g., Clarias) in their form and arrangement, and it seems probable that the Placodermi were annectent forms between the physostome Teleostei and the Ganoidei.
7. The Acanthodidae, on the other hand, seem to have connected the Ganoidei with the Elasmobranchii. The scales of these fishes of the Devonian and Carboniferous formations are very small, and similar to shagreen; spines, resembling the dermal defences of the Elasmobranchii, are placed in front of more, or fewer, of the median and of the paired fins. The skull appears to have been unossified, and the pectoral arch seems to have consisted of a single bony hoop.
The Pycnodontidae, which are commonly grouped among the Ganoids, are fishes with much-compressed bodies, like the John Dory or the Filefishes, covered with large rhomboidal enamelled scales, from which bony ridges projected internally, and were imbedded in the integument. The notochord is persistent, but the neural arches and the ribs are ossified. The proximal ends of the ribs, imbedded in the sheath of the notochord, are but little expanded in the more ancient members of the group, while, in the more modern species, they enlarge, and at length unite by serrated sutures, giving rise to spurious vertebrae. The skull is high and narrow, as in Balistes; the premaxillae are small, and there are no teeth in the maxillae, but several longitudinal series of crushing teeth (the vomer and parasphenoid ?) are attached to the base of the skull. These bite between the rami of the mandible, which are also armed with several rows of similar teeth. The teeth of the Pycnodonts have no vertical successors. The pectoral fins are small, the ventral, obsolete. The Pycnodonts are all extinct, but existed, formerly, for a very long period of time-their fossil remains occurring in rocks from the Carboniferous to the older Tertiary formations, inclusively. They present curious features of resemblance to the plectognath Teleostei.
The remains of Ganoid fishes began to appear in the Upper Silurian rocks at the same time as those of the Elasmobranchii, with which they constitute the oldest Vertebrata Fauna; they abound in the Devonian formation, and constitute, with the Elasmobranchii, the whole of the Palaeozoic Fish Fauna. We are in ignorance of the true affinities of Tharsis and Thrissops, and of the Hoplopleuridae; but unless some, or all, of these are Teleosteans, Ganoids and Elasmobranchs, alone, constitute the Fish Fauna of the Mesozoic formations, as far as the bottom of the Cretaceous series.
Support our developers




