The Pelvis of a bird
The pelvis of a bird (Fig. 86), is remarkable for the great elongation, both anteriorly and posteriorly, of the iliac bones (Il.), which unite with the whole length of the edges of the sacrum (Sm.) and even extend forward over the posterior ribs of the dorsal region. Below, each iliac bone forms a wide arch over the upper part of the acetabulum (Am.), the centre of which is always closed by fibrous tissue, so that, in the dry skeleton, the bottom of the acetabulum is perforated by a wide foramen. An articular surface on the ilium, on which the great trochanter of the femur plays, is called the antitrochanter. In all ordinary birds, the ischium (Fig. 86, Is.), which broadens toward its hinder end, extends back, nearly parallel with the hinder part of the ilium, and is united with it by ossification, posteriorly. The iliosciatic interval is thus converted into a foramen. The pubis (Pb.) enters, by its dorsal or acetabular end, into the formation of the acetabulum, and then passes backward and downward as a comparatively slender, curved, bone, nearly parallel with the ischium. It is united with its fellow only by fibrous tissue. Neither the ischia, nor the pubes, unite directly with the sacrum. Very few birds present any important deviation from this structure of the pelvis. In Titnamvus, Casuarius, Dromoeus, Apteryx, Dinornis, the ischium is not united with the backward extension of the ilium by bone.In Rhea, the ischia unite with one another beneath the vertebral column, and the vertebrae in this region are very slender and imperfectly ossified. In Struthio, alone, among birds, do the pubes unite in a median ventral symphysis.
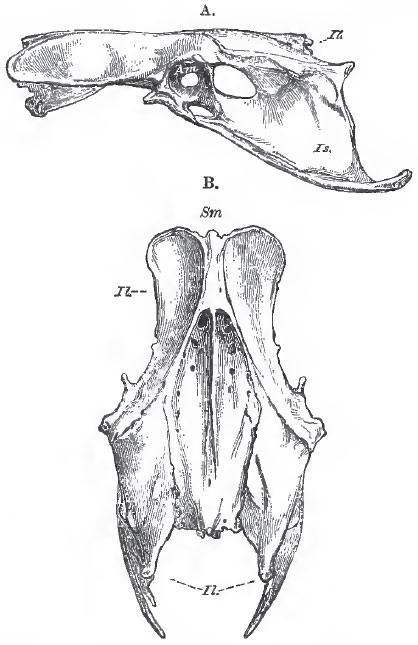 |
| Fig. 86. - A, lateral; and B, dorsal, view of the pelvis of a Fowl. Sm., sacrum; Il., illum Is., ischium; Pb., pubis; Am., acetabulum. |
The upper articular head of the femur is rounded, and its axis is almost at right angles with the body of the bone; a structure which is not found in ordinary Reptilia, but exists in the Iguanodon and other Ornithoscelida. The shaft is relatively short and thick, and the two terminal condyles are large and elongated antero-posteriorly. A prominent ridge, which plays between the proximal ends of the tibia and the fibula, is apparent upon the posterior and inferior surface of the outer condyle. A similar ridge is faintly developed in some Lacertilia, and is well marked in the Dinosaurian reptiles. A patella is usually present, but it is sometimes absent, and may be double.
The fibula of birds is always imperfect, ending in a mere style below. Generally it is decidedly shorter than the tibia, but it has the same length as that bone in some Penguins. The tibia, or rather tibio-tarsus, is a highly-characteristic bone. Its proximal end is expanded and produced anteriorly, into a great cnemial process (which may be variously subdivided) as in Dinosauria. The distal end is terminated by a wellmarked, pulley-like, articular surface which is inclined somewhat forward as well as downward. Not unfrequently there is an oblique bar of bone on the anterior face, just above the pulley, beneath which the long extensor tendons pass.
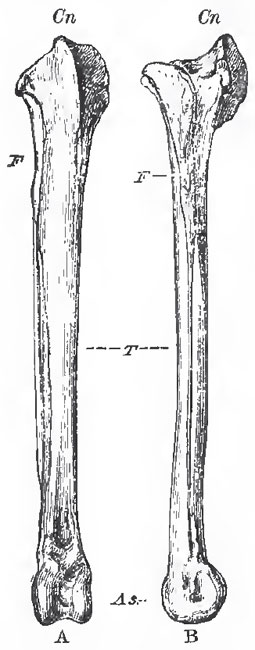 |
| Fig. 87. - The right tibia and fibula of a Fowl. A, front view: B. external lateral view. T., tibia; F., fibula; Cn., cremial process; As., astragalus. |
In all birds, even in Archoeopteryx, the fifth digit of the pes remains undeveloped; and the second, third, and fourth metatarsals are anchylosed together, and, by their proximal ends, with a bone, which is a distinct cartilage in the foetus, and represents the distal division of the tarsus. Thus a tarsometatarsus is formed. The distal ends of the metatarsals remain separate, and offer convex articular surfaces to the proximal phalanges of the digits.
In the Penguins, large apertures lie between the several metatarsals of the adult tarso-metatarsus; and, in other birds, more or less considerable passages persist between the middle and the lateral metatarsals proximally, and the middle and the outer, distally. In most birds, the middle metatarsal does not remain parallel with the others, but its proximal end inclines a little backward, and its distal end a little forward. Hence the two apertures on each side of its proximal end may lie at the bottom of a fossa, or run into one, in front, while they remain distinct behind.
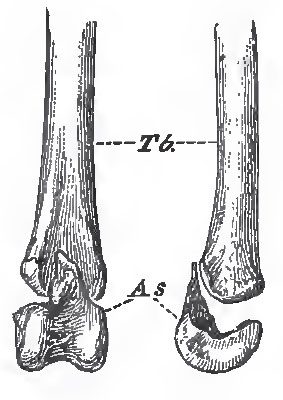 |
| Fig. 88. - The distal end of the left tibia (Tb.) with the astragalus (As.) detached, of a young Fowl. Viewed from in front, and from the outer side. |
When a hallux exists, its metatarsal bone is usually incomplete above, and is united by ligament to the inner, or the posterior, surface of the tarso-metatarsus. In the Frigate-bird (Phaethon) and in Steatornis, the hallucal metatarsal is remarkably long. The genus Phaethon stands alone, so far as I know, in having the hallucal metatarsal anchylosed with the others.
In many of the Alectoromorphae, a spur (calcar), consisting of a bony core ensheatlied in horn, is developed on the inner side of the metatarsus, and becomes anchylosed with the metatarsal of the second digit. In a few birds, similar spurs (Falamedea), or osseous excrescences (Pezophaps), are developed in relation with the metacarpus.
The normal number of the pedal phalanges in birds is (as in ordinary Lacertilia) 2, 3, 4, 5, reckoning from the hallux to the fourth digit. Among the few birds which constitute exceptions to the rule are the Swifts, in which the third and fourth toe have only three phalanges each (3, 3, 3, 3), and the Goatsuckers and the Sand-grouse, in which the fourth toe only has the number thus reduced (2, 3, 4, 3).
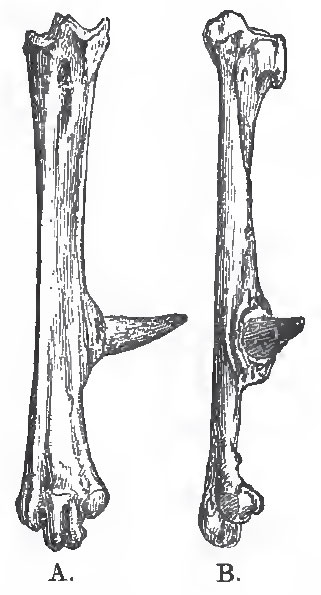 |
| Fig. 89. - The right tarso-metatarsus of a Fowl, consisting of three digits, ii., iii., iv., anchylosed with one another, and with the osseous core of the spur. A., front aspect; B., inner aspect. |
In most four-toed birds the hallux is turned more or less completely backward, and the other three digits forward. But in many Aetomorphae (especially the Owls), the outer toe can be turned outward, or even backward, at will. And in the Parrots, Toucans, Cuckoos, Woodpeckers, and other so-called "scansorial" birds, the outer toe is permanently reversed. Under these circumstances the distal end of the outer metatarsal may be divided into two distinct articular surfaces. In the Trogons, there are two toes in front and two behind, as in the Parrots, but it is the second toe which is turned backward. Lastly, in the Swifts, the Dysporomorphae and the Spheniscomorphae, the hallux is directed more or less forward, so that all four toes are turned to the front.
As a general rule, the osseous tissue of birds is remarkably dense and hard. Before hatching, the bones are solid and filled with vascular medulla; but, after birth, more or fewer of the bones are always excavated by prolongations of cavities containing air, which lie in their neighborhood. Such aircavities are always found in the skull, in connection with the nasal and auditory passages, and they may extend through all parts of the skull, with the exception of the jugal arch. In many birds, such as the Apteryx, Penguins, Divers, Gulls, and the smaller song-birds, no other bones than those of the skull are pneumatic; but, in most birds, the air-sacs of the lungs send off prolongations into the bones of the rest of the skeleton, and thus the whole skeleton in some cases (as in the Hornbills) becomes pneumatic. It is proper to remark that the amount of pneumaticity of the bones by no means follows the development of the power of flight. In the Ostrich, for example, the bones are far more extensively pneumatic than in the Gull.
In some cases, prolongations of the air-sacs extend beneath the integument.
Support our developers




