Fungus
List of Plant Organisms
» Tree
» Herbs
» Bush/Shrub
» Grass
» Vines
» Fern
» Moss
» Fern Ally
» Algae
» Fungi
A fungus is a eukaryotic organism that is a member of the kingdom Fungi. The fungi are a monophyletic group, also called the Eumycota ("true fungi" or eumycetes), that is phylogenetically distinct from the morphologically similar slime molds (myxomycetes) and water molds (oomycetes). The fungi are heterotrophic organisms possessing a chitinous cell wall, with the majority of fungal species growing as multicellular filaments called hyphae forming a mycelium; some fungal species also grow as single cells. Sexual and asexual reproduction of the fungi is commonly via spores, often produced on specialized structures or in fruiting bodies. Some species have lost the ability to form reproductive structures, and propagate solely by vegetative growth. Yeasts, molds, and mushrooms are examples of fungi. The fungi are more closely related to animals than plants, yet the discipline of biology devoted to the study of fungi, known as mycology, often falls under a branch of botany.» Tree
» Herbs
» Bush/Shrub
» Grass
» Vines
» Fern
» Moss
» Fern Ally
» Algae
» Fungi
Occurring worldwide, most fungi are largely invisible to the naked eye, living for the most part in soil, dead matter, and as symbionts of plants, animals, or other fungi. They perform an essential role in all ecosystems in decomposing organic matter and are indispensable in nutrient cycling and exchange.

Clockwise from top left: Amanita muscaria, a basidiomycete; Sarcoscypha coccinea, an ascomycete; black bread mold, a zygomycete; a chytrid; a Penicillium conidiophore.
Etymology and definition
The English word fungus is directly adopted from the Latin fungus, meaning "mushroom", used in Horace and Pliny. This in turn is derived from the Greek word sphongos/σφογγος ("sponge"), referring to the macroscopic structures and morphology of some mushrooms and molds and also used in other languages (e.g., the German Schwamm ("sponge") or Schwammerl for some types of mushroom).Characteristics
The fungi have a range of features defining the fungal kingdom, some of which are shared with other organisms while others are unique to the fungi. Shared features:» With eukaryotes: All fungi are eukaryotic, containing membrane-bound nuclei with chromosomes. Fungal cells contain membrane-bound cytoplasmic organelles, DNA with noncoding regions called introns, sterol-containing membranes, and ribosomes of the 80S type. Fungi have a characteristic range of soluble carbohydrates and storage compounds, including mannitol and other sugar alcohols, trehalose and glycogen the latter of which is also found in animals.
» With animals: Fungi lack chloroplasts and are heterotrophic organisms, requiring preformed organic compounds as energy sources and also as carbon skeletons for organic synthesis.
» With plants: Fungi possess a cell wall. They reproduce by both sexual and asexual means, and like some basal plant groups, such as ferns and mosses produce spores. Similar to mosses and algae, fungi typically have haploid nuclei.
» With prokaryotes: As in some bacteria, biosynthesis of the amino acid, L-lysine, is via the α-aminoadipate pathway.
» Fungi typically grow as hyphae, which extend at their tips. This apical growth form is shared with the morphologically similar oomycetes and in contrast with other filamentous organisms, like filamentous green algae, which grow by repeated cell divisions within a chain of cells (intercalary growth).
» Some fungi grow as single-celled yeasts which reproduce by budding, and some dimorphic fungi can switch between a yeast phase and a hyphal phase in response to environmental conditions.
» The fungal cell wall contains glucans also found in plants, but also chitin not found in the Plant kingdom, but in some animals. In contrast to plants and the oomycetes, fungal cell walls do not contain cellulose.
» Fungal hyphae may have several nuclei within each hyphal compartment, and many budding yeasts are diploid.
Diversity
Fungi have a worldwide distribution, and grow in a wide range of habitats, including deserts, hypersaline environments, the deep sea, on rocks, and in extremely low and high temperatures. They have been shown to be able to survive the intense UV and cosmic radiation encountered during space travel. Most fungi grow in terrestrial environments, but several species live part or all of their lives in aquatic habitats. For example, the chytrid fungus Batrachochytrium dendrobatidis—responsible for a worldwide decline in amphibian populations—spends part of its life cycle as motile zoospore, enabling it to propel itself through water and penetrate the skin of an amphibian host. Fungi along with bacteria are the primary decomposers of organic matter in most if not all terrestrial ecosystems worldwide. Based on observations of the ratio of the number of fungal species to the number of plant species in some environments, the fungal kingdom has been estimated to contain about 1.5 million species. Around 70,000 fungal species have been formally described by taxonomists, but the true dimension of fungal diversity is still unknown. Most fungi grow as thread-like filaments called hyphae, which form mycelia, while others grow as single cells. Until recently, many fungal species were described based mainly on morphological characteristics, such as the size and shape of spores or fruiting structures, and biological species concepts. The application of molecular tools, such as DNA sequencing and phylogenetic analysis, to study fungal diversity has greatly enhanced the resolution and added robustness to estimates of genetic diversity within various taxonomic groups.Morphology
Microscopic structures

Mold covering a decaying peach over a period of six days. The frames were taken approximately 12 hours apart.
Many fungi grow as thread-like filamentous microscopic structures called hyphae, and an assemblage of intertwined and interconnected hyphae is called a mycelium. Hyphae can be septate, i.e., divided into hyphal compartments separated by a septum, each compartment containing one or more nuclei or can be coenocytic, i.e., lacking hyphal compartmentalization. However, septa have pores, such as the doliporus in the basidiomycetes that allow cytoplasm, organelles, and sometimes nuclei to pass through. Coenocytic hyphae are essentially multinucleate supercells. In some cases, fungi have developed specialized structures for nutrient uptake from living hosts; examples include haustoria in plant-parasitic fungi of nearly all divisions, and arbuscules of several mycorrhizal fungi, which penetrate into the host cells for nutrient uptake by the fungus.
Macroscopic structures
Fungal mycelia can become visible macroscopically, for example, as concentric rings on various surfaces, such as damp walls, and on other substrates, such as spoilt food (see figure), and are commonly and generically called mould (American spelling, mold); fungal mycelia grown on solid agar media in laboratory petri dishes are usually referred to as colonies, with many species exhibiting characteristic macroscopic growth morphologies and colours, due to spores or pigmentation.Specialized fungal structures important in sexual reproduction are the apothecia, perithecia, and cleistothecia in the ascomycetes, and the fruiting bodies of the basidiomycetes, and a few ascomycetes. These reproductive structures can sometimes grow very large, and are well known as mushrooms.
Growth and physiology
Growth of fungi as filamentous hyphae on or in solid substrates or single cells in aquatic environments is adapted to efficient extraction of nutrients from these environments, because these growth forms have high surface area to volume ratios. Fungal hyphae are specifically adapted to growth on solid surfaces and within substrates, and can exert large penetrative mechanical forces. The plant pathogen, Magnaporthe grisea, forms a structure called an appressorium specifically designed for penetration of plant tissues. The pressure generated by the appressorium, which is directed against the plant epidermis, can exceed 8 MPa (80 bars). A similar method is employed by the filamentous fungus Paecilomyces lilacinus to penetrate the eggs of plant-parasitic nematodes. The generation of these mechanical pressures is the result of an interplay between physiological processes to increase intracellular turgor by production of osmolytes such as glycerol, and the morphology of the appressorium. These adaptations in morphology are complemented by hydrolytic enzymes secreted into the environment for digestion of large organic molecules, such as polysaccharides, proteins, lipids, and other organic substrates into smaller molecules. These molecules are then absorbed as nutrients into the fungal cells.Traditionally, the fungi are considered heterotrophs, organisms that rely solely on carbon fixed by other organisms for metabolism. Fungi have evolved a remarkable metabolic versatility that allows many of them to use a large variety of organic substrates for growth, including simple compounds as nitrate, ammonia, acetate, or ethanol. Recent research raises the possibility that some fungi utilize the pigment melanin to extract energy from ionizing radiation, such as gamma radiation for "radiotrophic" growth. It has been proposed that this process might bear some similarity to photosynthesis in plants, but detailed biochemical data supporting the existence of this hypothetical pathway are presently lacking.
Reproduction
Reproduction of fungi is complex, reflecting the heterogeneity in lifestyles and genetic make up within this group of organisms. Many fungi reproduce either sexually or asexually, depending on conditions in the environment. These conditions trigger genetically determined developmental programs leading to the expression of specialized structures for sexual or asexual reproduction. These structures aid both reproduction and efficient dissemination of spores or spore-containing propagules.Asexual reproduction
Asexual reproduction via vegetative spores or through mycelial fragmentation is common in many fungal species and allows more rapid dispersal than sexual reproduction.
Fungi on a fence post near Orosí, Costa Rica.
Sexual reproduction
Sexual reproduction with meiosis exists in all fungal phyla, except the Deuteromycota. It differs in many aspects from sexual reproduction in animals or plants. Many differences also exist between fungal groups and have been used to discriminate fungal clades and species based on morphological differences in sexual structures and reproductive strategies. Experimental crosses between fungal isolates can also be used to identify species based on biological species concepts. The major fungal clades have initially been delineated based on the morphology of their sexual structures and spores; for example, the spore-containing structures, asci and basidia, can be used in the identification of ascomycetes and basidiomycetes, respectively. Many fungal species have elaborate vegetative incompatibility systems that allow mating only between individuals of opposite mating type, while others can mate and sexually reproduce with any other individual or itself. Species of the former mating system are called heterothallic, and of the latter homothallic.Most fungi have both a haploid and diploid stage in their life cycles. In all sexually reproducing fungi, compatible individuals combine by cell fusion of vegetative hyphae by anastomosis, required for the initiation of the sexual cycle. Ascomycetes and basidiomycetes go through a dikaryotic stage, in which the nuclei inherited from the two parents do not fuse immediately after cell fusion, but remain separate in the hyphal cells (see heterokaryosis).
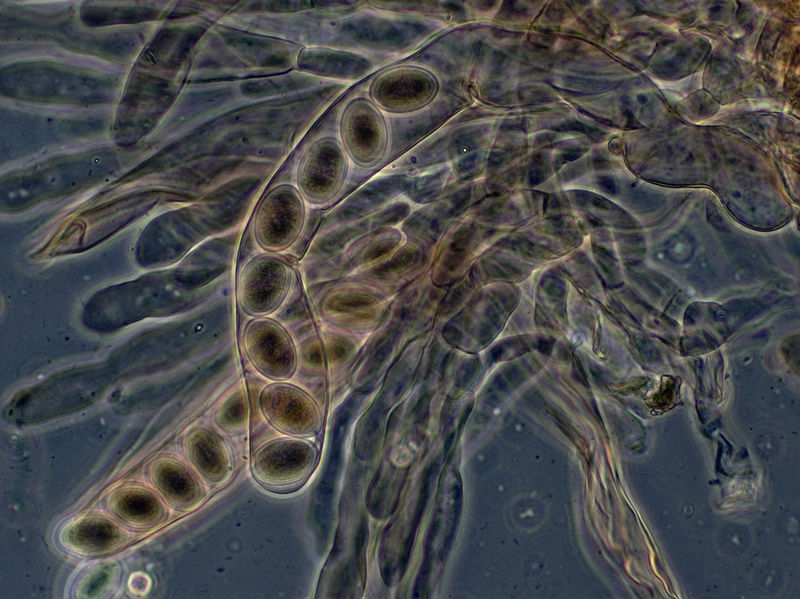
The 8-spored asci of Morchella elata, viewed with phase contrast microscopy.
Sexual reproduction in basidiomycetes is similar to that of the ascomycetes. Compatible haploid hyphae fuse to produce a dikaryotic mycelium. However, the dikaryotic phase is more extensive in the basidiomycetes, in many cases also present in the vegetatively growing mycelium. A specialized anatomical structure, called a clamp connection, is formed at each hyphal septum.
In zygomycetes, haploid hyphae of two individuals fuse, forming a zygote, which develops into a zygospore. When the zygospore germinates, it quickly undergoes meiosis, generating new haploid hyphae, which in turn may form asexual sporangiospores. These sporangiospores are means of rapid dispersal of the fungus and germinate into new genetically identical haploid fungal colonies, able to mate and undergo another sexual cycle followed by the generation of new zygospores, thus completing the lifecycle.
Spore dispersal
Both asexual and sexual spores or sporangiospores of many fungal species are actively dispersed by forcible ejection from their reproductive structures. This ejection ensures exit of the spores from the reproductive structures as well as travelling through the air over long distances. Many fungi thereby possess specialized mechanical and physiological mechanisms as well as spore-surface structures, such as hydrophobins, for spore ejection. These mechanisms include, for example, forcible discharge of ascospores enabled by the structure of the ascus and accumulation of osmolytes in the fluids of the ascus that lead to explosive discharge of the ascospores into the air. The forcible discharge of single spores termed ballistospores involves formation of a small drop of water (Buller's drop), which upon contact with the spore leads to its projectile release with an initial acceleration of more than 10,000 g. Other fungi rely on alternative mechanisms for spore release, such as external mechanical forces, exemplified by puffballs. Attracting insects, such as flies, to fruiting structures, by virtue of their having lively colours and a putrid odour, for dispersal of fungal spores is yet another strategy, most prominently used by the stinkhorns.Other sexual processes
Besides regular sexual reproduction with meiosis, some fungal species may exchange genetic material via parasexual processes, initiated by anastomosis between hyphae and plasmogamy of fungal cells. The frequency and relative importance of parasexual events is unclear and may be lower than other sexual processes. However, it is known to play a role in intraspecific hybridization and is also likely required for hybridization between fungal species, which has been associated with major events in fungal evolution.Phylogeny and classification
For a long time taxonomists considered fungi to be members of the Plant Kingdom. This early classification was based mainly on similarities in lifestyle: both fungi and plant are mainly sessile, have similarities in general morphology and growth habitat (like plants, fungi often grow in soil, in the case of mushrooms forming conspicuous fruiting bodies, which sometimes bear resemblance to plants such as mosses). Moreover, both groups possess a cell wall, which is absent in the Animal Kingdom. However, the fungi are now considered a separate kingdom, distinct from both plants and animals, from which they appear to have diverged approximately one billion years ago. Many studies have identified several distinct morphological, biochemical, and genetic features in the Fungi, clearly delineating this group from the other kingdoms.
The parasitic mushroom Oudemansiella nocturnum, infesting the wood of a tree.
Physiological and morphological traits
Similar to animals and unlike most plants, fungi lack the capacity to synthesize organic carbon by chlorophyll-based photosynthesis; whereas plants store the reduced carbon as starch, fungi, like animals and some bacteria, use glycogen for storage of carbohydrates. A major component of the cell wall in many fungal species is the nitrogen-containing carbohydrate, chitin, also present in some animals, such as the insects and crustaceans, while the plant cell wall consists chiefly of the carbohydrate cellulose. The defining and unique characteristics of fungal cells include growth as hyphae, which are microscopic filaments of between 2-10 microns in diameter and up to several centimetres in length, and which combined form the fungal mycelium. Some fungi, such as yeasts, grow as single ovoid cells, similar to unicellular algae and the protists.Some characteristics shared between plants and fungi include the presence of vacuoles in the cell, and a similar pathway in the biosynthesis of terpenes using mevalonic acid and pyrophosphate as biochemical precursors; plants however use an additional terpene biosynthesis pathway in the chloroplasts that is apparently absent in fungi. Ancestral traits shared among members of the fungi include chitinous cell walls and heterotrophy by absorption. A further characteristic of the fungi that is absent from other eukaryotes, and shared only with some bacteria, is the biosynthesis of the amino acid, L-lysine, via the α-aminoadipate pathway.
Similar to plants, fungi produce a plethora of secondary metabolites functioning as defensive compounds or for niche adaptation; however, biochemical pathways for the synthesis of similar or even identical compounds often differ markedly between fungi and plants.
Evolutionary history
The first organisms having features typical of fungi date to 1,200 million years ago, the Proterozoic. However, fungal fossils do not become common and uncontroversial until the early Devonian, when they are abundant in the Rhynie chert.Even though traditionally included in many botany curricula and textbooks, fungi are now thought to be more closely related to animals than to plants and are placed with the animals in the monophyletic group of opisthokonts. For much of the Paleozoic Era, the fungi appear to have been aquatic, and consisted of organisms similar to the extant Chytrids in having flagellum-bearing spores. The early fossil record of the fungi is fragmentary, to say the least. The fungi probably colonized the land during the Cambrian, long before land plants. All modern classes of fungi were present in the Late Carboniferous (Pennsylvanian Epoch). For some time after the Permian-Triassic extinction event, a fungal spike, originally thought to be an extraordinary abundance of fungal spores in sediments formed shortly after this event, suggested that they were the dominant life form during this period—nearly 100% of the fossil record available from this period. However, the relative proportion of fungal spores relative to spores formed by algal species is difficult to assess, the spike did not appear worldwide, and in many places it did not fall on the Permian-Triassic boundary.
There is no unique generally accepted system at the higher taxonomic levels and there are constant name changes at every level, from species upwards. However, efforts among fungal researchers are now underway to establish and encourage usage of a unified and more consistent nomenclature. Fungal species can also have multiple scientific names depending on its life cycle and mode (sexual or asexual) of reproduction. Web sites such as Index Fungorum and ITIS define preferred up-to-date names (with cross-references to older synonyms), but do not always agree with each other.
Cladogram
A cladogram depicts the phylogenetic relationships between several groups of organisms in a tree-like diagram. The current classification of Kingdom Fungi recognizes seven phyla, two of which—the Ascomycota and the Basidiomycota—are contained within a branch representing subkingdom Dikarya.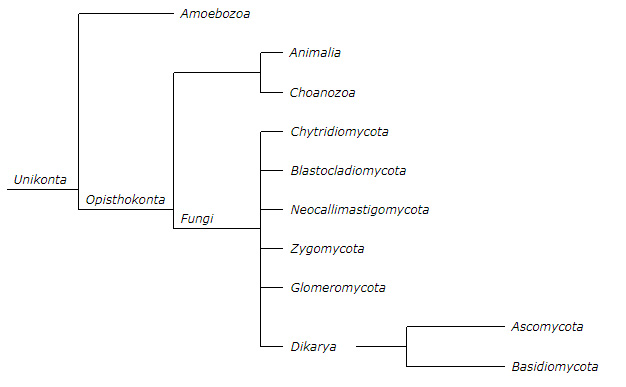
Taxonomic groups
The major divisions (phyla) of fungi have been classified based mainly on the characteristics of their sexual reproductive structures. Currently, seven fungal divisions are proposed:Chytridiomycota
The Chytridiomycota are commonly known as chytrids. These fungi are ubiquitous with a worldwide distribution. Chytrids produce zoospores that are capable of active movement through aqueous phases with a single flagellum, leading some early taxonomists to classify them as protists. Molecular phylogenies, inferred from rRNA sequences in ribosomes, suggest that the Chytrids are a basal fungal group divergent from the other fungal divisions, consisting of four major clades with some evidence for paraphyly or possibly polyphyly.Blastocladiomycota
The Blastocladiomycota were previously considered a taxonomic clade within the Chytridiomycota. Recent molecular data and ultrastructural characteristics, however, place the Blastocladiomycota as a sister clade to the Zygomycota, Glomeromycota, and Dikarya (Ascomycota and Basiomycota). The blastocladiomycetes are fungi that are saprotrophs and parasites of all eukaryotic groups and undergo sporic meiosis unlike their close relatives, the chytrids, which mostly exhibit zygotic meiosis.Neocallimastigomycota

Arbuscular mycorrhiza seen under microscope. Flax root cortical cells containing paired arbuscules.
The Neocallimastigomycota were earlier placed in the phylum Chytridomycota. Members of this small phylum are anaerobic organisms, living in the digestive system of larger herbivorous mammals and possibly in other terrestrial and aquatic environments. They lack mitochondria but contain hydrogenosomes of mitochondrial origin. As the related chrytrids, neocallimastigomycetes form zoospores that are posteriorly uniflagellate or polyflagellate.
Zygomycota
The Zygomycota contain the taxa, Zygomycetes and Trichomycetes, and reproduce sexually with meiospores called zygospores and asexually with sporangiospores. Black bread mold (Rhizopus stolonifer) is a common species that belongs to this group; another is Pilobolus, which is capable of ejecting spores several meters through the air. Medically relevant genera include Mucor, Rhizomucor, and Rhizopus. Molecular phylogenetic investigation has shown the Zygomycota to be a polyphyletic phylum with evidence of paraphyly within this taxonomic group.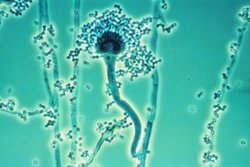
Conidiophores of molds of the genus Aspergillus, an ascomycete, seen under microscope.
Glomeromycota
Members of the Glomeromycota are fungi forming arbuscular mycorrhizae with higher plants. Only one species has been observed forming zygospores; all other species solely reproduce asexually. The symbiotic association between the Glomeromycota and plants is ancient, with evidence dating to 400 million years ago.Ascomycota
The Ascomycota, commonly known as sac fungi or ascomycetes, constitute the largest taxonomic group within the Eumycota. These fungi form meiotic spores called ascospores, which are enclosed in a special sac-like structure called an ascus. This division includes morels, a few mushrooms and truffles, single-celled yeasts (e.g., of the genera Saccharomyces, Kluyveromyces, Pichia, and Candida), and many filamentous fungi living as saprotrophs, parasites, and mutualistic symbionts. Prominent and important genera of filamentous ascomycetes include Aspergillus, Penicillium, Fusarium, and Claviceps. Many ascomycetes species have only been observed undergoing asexual reproduction (called anamorphic species), but analysis of molecular data has often been able to identify their closest teleomorphs in the Ascomycota. Because the products of meiosis are retained within the sac-like ascus, several ascomycetes have been used for elucidating principles of genetics and heredity (e.g. Neurospora crassa).Basidiomycota
Members of the Basidiomycota, commonly known as the club fungi or basidiomycetes, produce meiospores called basidiospores on club-like stalks called basidia. Most common mushrooms belong to this group, as well as rust and smut fungi, which are major pathogens of grains. Other important Basidiomycetes include the maize pathogen Ustilago maydis, human commensal species of the genus Malassezia, and the opportunistic human pathogen, Cryptococcus neoformans.Phylogenetic relationships with other fungus-like organisms
Because of some similarities in morphology and lifestyle, the slime molds (myxomycetes) and water molds (oomycetes) were formerly classified in the kingdom Fungi. Unlike true fungi, however, the cell walls of these organisms contain cellulose and lack chitin. Slime molds are unikonts like fungi, but are grouped in the Amoebozoa. Water molds are diploid bikonts, grouped in the Chromalveolate kingdom. Neither water molds nor slime molds are closely related to the true fungi, and, therefore, taxonomists no longer group them in the kingdom Fungi. Nonetheless, studies of the oomycetes and myxomycetes are still often included in mycology textbooks and primary research literature.It has been suggested that the nucleariids, currently grouped in the Choanozoa, may be a sister group to the eumycete clade, and as such could be included in an expanded fungal kingdom.
Ecology
Although often inconspicuous, fungi occur in every environment on Earth and play very important roles in most ecosystems. Along with bacteria, fungi are the major decomposers in most terrestrial (and some aquatic) ecosystems, and therefore play a critical role in biogeochemical cycles and in many food webs. As decomposers, they play an indispensable role in nutrient cycling, especially as saprotrophs and symbionts, degrading organic matter to inorganic molecules, which can then re-enter anabolic metabolic pathways in plants or other organisms.Symbiosis
Many fungi have important symbiotic relationships with organisms from most if not all Kingdoms. These interactions can be mutualistic or antagonistic in nature, or in case of commensal fungi are of no apparent benefit or detriment to the host.
Polypores growing on a tree in Borneo
With plants
Mycorrhizal symbiosis between plants and fungi is one of the most well-known plant-fungus associations and is of significant importance for plant growth and persistence in many ecosystems; over 90% of all plant species engage in some kind of mycorrhizal relationship with fungi and are dependent upon this relationship for survival. The mycorrhizal symbiosis is ancient, dating to at least 400 million years ago. It often increases the plant's uptake of inorganic compounds, such as nitrate and phosphate from soils having low concentrations of these key plant nutrients. In some mycorrhizal associations, the fungal partners may mediate plant-to-plant transfer of carbohydrates and other nutrients. Such mycorrhizal communities are called "common mycorrhizal networks".With algae and cyanobacteria
Lichens are formed by a symbiotic relationship between algae or cyanobacteria (referred to in lichens as "photobionts") and fungi (mostly various species of ascomycetes and a few basidiomycetes), in which individual photobiont cells are embedded in a tissue formed by the fungus. As in mycorrhizas, the photobiont provides sugars and other carbohydrates, while the fungus provides minerals and water.
The lichen Lobaria pulmonaria, a symbiosis of fungal, algal, and cyanobacterial species.
With insects
Many insects also engage in mutualistic relationships with various types of fungi. Several groups of ants cultivate fungi in the order Agaricales as their primary food source, while ambrosia beetles cultivate various species of fungi in the bark of trees that they infest. Termites on the African Savannah are also known to cultivate fungi.As pathogens and parasites
However, many fungi are parasites on plants, animals (including humans), and other fungi. Serious fungal pathogens of many cultivated plants causing extensive damage and losses to agriculture and forestry include the rice blast fungus Magnaporthe oryzae, tree pathogens such as Ophiostoma ulmi and Ophiostoma novo-ulmi causing Dutch elm disease, and Cryphonectria parasitica responsible for chestnut blight, and plant-pathogenic fungi in the genera Fusarium, Ustilago, Alternaria, and Cochliobolus. Some fungi are predators of nematodes, which they capture using an array of specialized structures, such as constricting rings or adhesive nets.Fungi are the causal agents of a variety of serious diseases in humans, several of which may be fatal if not treated properly. These include aspergilloses, candidoses, coccidioidomycosis, cryptococcosis, histoplasmosis, a variety of mycetomas, and paracoccidioidomycosis. Furthermore, persons with immuno-deficiencies are particularly susceptible to disease by fungal genera such as Aspergillus, Candida, Cryptoccocus, Histoplasma, and Pneumocystis. Other fungi can attack eyes, nails, hair, and especially skin, the so-called dermatophytic fungi and keratinophilic fungi, and cause a variety of conditions, of which ringworms such as athlete’s foot are common. Fungal spores are also a major cause of allergies, and a wide range of fungi from different taxonomic groups can evoke allergic reactions in some people.
Importance for human use
Human use of fungi for food preparation or preservation and other purposes is extensive and has a long history: yeasts are required for fermentation of beer, wine and bread, other fungal species are used in the production of soy sauce and tempeh. Mushroom farming and mushroom gathering are large industries in many countries. Many fungi are producers of antibiotics, including β-lactam antibiotics such as penicillin and cephalosporin. Widespread use of these antibiotics for the treatment of bacterial diseases, such as tuberculosis, syphilis, leprosy, and many others began in the early 20th century and continues to play a major part in anti-bacterial chemotherapy. The study of the historical uses and sociological impact of fungi is known as ethnomycology.Cultured foods
Baker's yeast or Saccharomyces cerevisiae, a single-cell fungus, is used in the baking of bread and other wheat-based products, such as pizza and dumplings.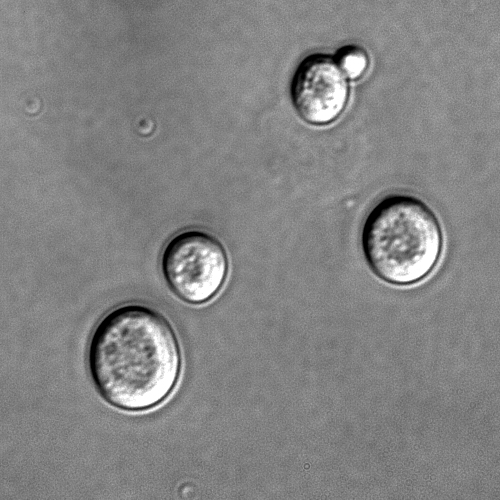
Saccharomyces cerevisiae cells shown with DIC microscopy.
Other human uses
Fungi are also used extensively to produce industrial chemicals like citric, gluconic, lactic, and malic acids, antibiotics, and even to make stonewashed jeans. Fungi are also sources of industrial enzymes, such as lipases used in biological detergents, amylases, cellulases, invertases, proteases and xylanases. Several fungal species are ingested for their psychedelic properties, both recreationally and religiously (see main article, Psilocybin mushrooms).Mycotoxins
Many fungi produce biologically active compounds, several of which are toxic and are therefore called mycotoxins. Of particular relevance to humans are those mycotoxins that are produced by molds causing food spoilage and poisonous mushrooms (see below). Particularly infamous are the aflatoxins, which are insidious liver toxins and highly carcinogenic metabolites produced by Aspergillus species often growing in or on grains and nuts consumed by humans, and the lethal amatoxins produced by mushrooms of the genus Amanita. Other notable mycotoxins include ochratoxins, patulin, ergot alkaloids, and trichothecenes and fumonisins, all of which have significant impact on human food supplies or animal livestock.Mycotoxins belong to the group of compounds known as secondary metabolites (or natural products). Originally, these compounds had been thought to be mere byproducts of primary metabolism, hence the name "secondary" metabolites. However, recent research has shown the existence of biochemical pathways solely for the purpose of producing mycotoxins and other natural products in fungi. Mycotoxins provide a number of fitness benefits to the fungi that produce them in terms of physiological adaptation, competition with other microbes and fungi, and protection from consumption (fungivory). These fitness benefits and the existence of dedicated biosynthetic pathways for mycotoxin production suggest that the mycotoxins are important for fungal persistence and survival.
Edible and poisonous fungi
Some of the best known types of fungi are the edible and the poisonous mushrooms. Many species are commercially raised, but others must be harvested from the wild. Agaricus bisporus, sold as button mushrooms when small or Portobello mushrooms when larger, are the most commonly eaten species, used in salads, soups, and many other dishes. Many Asian fungi are commercially grown and have gained in popularity in the West. They are often available fresh in grocery stores and markets, including straw mushrooms (Volvariella volvacea), oyster mushrooms (Pleurotus ostreatus), shiitakes (Lentinula edodes), and enokitake (Flammulina spp.).There are many more mushroom species that are harvested from the wild for personal consumption or commercial sale. Milk mushrooms, morels, chanterelles, truffles, black trumpets, and porcini mushrooms (Boletus edulis) (also known as king boletes) all demand a high price on the market. They are often used in gourmet dishes.
For certain types of cheeses, it is also a common practice to inoculate milk curds with fungal spores to promote the growth of specific species of mold that impart a unique flavor and texture to the cheese. This accounts for the blue colour in cheeses such as Stilton or Roquefort which is created using Penicillium roqueforti spores. Molds used in cheese production are usually non-toxic and are thus safe for human consumption; however, mycotoxins (e.g., aflatoxins, roquefortine C, patulin, or others) may accumulate due to fungal spoilage during cheese ripening or storage.
Many mushroom species are toxic to humans, with toxicities ranging from slight digestive problems or allergic reactions as well as hallucinations to severe organ failures and death. Some of the most deadly mushrooms belong to the genera Inocybe, Cortinarius, and most infamously, Amanita. The latter genus includes the destroying angel (A. virosa) and the death cap (A. phalloides), the most common cause of deadly mushroom poisoning. The false morel (Gyromitra esculenta) is considered a delicacy by some when cooked, yet can be highly toxic when eaten raw. Tricholoma equestre was considered edible until being implicated in some serious poisonings causing rhabdomyolysis.

Black Périgord Truffle (Tuber melanosporum), cut in half.
Fungi in the biological control of pests

Asian mushrooms, clockwise from left, enokitake, buna-shimeji, bunapi-shimeji, king oyster mushroom and shiitake.
Bioremediation

Stilton cheese veined with Penicillium roqueforti.
Notes and References
- Moore RT. (1980). "Taxonomic proposals for the classification of marine yeasts and other yeast-like fungi including the smuts". Botanica Marine 23: 361–73.
- These are the pronunciations listed first in most American dictionaries. See, for example, the Merriam-Webster Online entry. In British dictionaries, such as the Oxford Advanced Learner's Dictionary and Cambridge's English Pronouncing Dictionary, the pronunciations given for fungi are /ˈfʌŋgaɪ/ and /ˈfʌŋgi/. Funguses (/ˈfʌŋgəsəz/) is an alternative plural form.
- Simpson, DP. (1979). Cassell's Latin Dictionary (5 ed.). London: Cassell Ltd. p. 883.
- Alexopoulos CJ, Mims CW, Blackwell M. (1996). Introductory Mycology. John Wiley and Sons.
- Vaupotic T, Veranic P, Jenoe P, Plemenitas A. (2008). "Mitochondrial mediation of environmental osmolytes discrimination during osmoadaptation in the extremely halotolerant black yeast Hortaea werneckii". Fungal Genetics and Biology 45 (6): 994–1007. doi:10.1016/j.fgb.2008.01.006.
- Raghukumar C, Raghukumar S. (1998). "Barotolerance of fungi isolated from deep-sea sediments of the Indian Ocean". Aquatic Microbial Ecology 15: 153–63.
- Garvie LA, Knauth LP, Bungartz F, Klonowski S, Nash TH. (August 2008). "Life in extreme environments: survival strategy of the endolithic desert lichen Verrucaria rubrocincta". Naturwissenschaften 95 (8): 705–12.
- Sancho LG, de la Torre R, Horneck G, Ascaso C, de Los Rios A, Pintado A, Wierzchos J, Schuster M. (2007). "Lichens survive in space: results from the 2005 LICHENS experiment". Astrobiology 7 (3): 443–54.
- Brem FM, Lips KR (2008). "Batrachochytrium dendrobatidis infection patterns among Panamanian amphibian species, habitats and elevations during epizootic and enzootic stages". Diseases of Aquatic Organisms 81 (3): 189–202.
- Hawksworth DL. (2006). "The fungal dimension of biodiversity: magnitude, significance, and conservation". Mycological Research 95: 641–55.
- Mueller GM, Schmit JP. (2006). "Fungal biodiversity: what do we know? What can we predict?". Biodiversity and Conservation 16: 1–5.
- Meredith Blackwell; Rytas Vilgalys, and John W. Taylor (2005-02-14). "Eumycota: mushrooms, sac fungi, yeast, molds, rusts, smuts, etc.".
- Hibbett DS, et al. (2007). "A higher level phylogenetic classification of the Fungi". Mycological Research 111 (5): 509–47.
- Steenkamp ET, Wright J, Baldauf SL. (2006). The protistan origins of animals and fungi. Molecular Biology and Evolution 23 (1): 93-106.
- Stevens DA, Ichinomiya M, Koshi Y, Horiuchi H. (2006). "Escape of Candida from caspofungin inhibition at concentrations above the MIC (paradoxical effect) accomplished by increased cell wall chitin; evidence for β-1,6-glucan synthesis inhibition by caspofungin". Antimicrobial Agents and Chemotherapy 50: 3160–1.
- Chang, S-T; Miles PG. (2004). Mushrooms: Cultivation, Nutritional Value, Medicinal Effect and Environmental Impact.
- Howard RJ, Ferrari MA, Roach DH, Money NP. (1991). "Penetration of hard substrates by a fungus employing enormous turgor pressures". Proceedings of the National Academy of Sciences U S A. 88: 11281–4.
- Money, NP (1998). Mechanics of invasive fungal growth and the significance of turgor in plant infection. In: Molecular genetics of host-specific toxins in plant disease. Netherlands: Kluwer Academic Publishers. pp. 261–271.
- Wang ZY, Jenkinson JM, Holcombe LJ, Soanes DM, Veneault-Fourrey C, Bhambra GK, Talbot NJ. (2005). "The molecular biology of appressorium turgor generation by the rice blast fungus Magnaporthe grisea". Biochemical Society Transactions 33: 384–8.
- Pereira JL, Noronha EF, Miller RN, Franco OL. (2007). "Novel insights in the use of hydrolytic enzymes secreted by fungi with biotechnological potential". Letters in Applied Microbiology 44: 573–81.
- Schaller M, Borelli C, Korting HC, Hube B. (2007). "Hydrolytic enzymes as virulence factors of Candida albicans". Mycoses 48: 365–77.
- Farrar JF (1985). "Carbohydrate metabolism in biotrophic plant pathogens". Microbiological Sciences 2: 314–7.
- Marzluf GA. (1981). "Regulation of nitrogen metabolism and gene expression in fungi". Microbiological Reviews 45: 437–61.
- Heynes MJ (1994). "Regulatory circuits of the amdS gene of Aspergillus nidulans". Antonie Van Leeuwenhoek 65: 179–82.
- Dadachova E, Bryan RA, Huang X, Moadel T, Schweitzer AD, Aisen P, Nosanchuk JD, Casadevall A. (2007). "Ionizing radiation changes the electronic properties of melanin and enhances the growth of melanized fungi".
- Metzenberg RL, Glass NL. (1990). "Mating type and mating strategies in Neurospora". Bioessays 12: 53–9. doi:10.1002/bies.950120202.
- Sitte P, Ziegler H, Ehrendorfer F. (1991). Strasburger Lehrbuch der Botanik (Textbook of Botany) (33 ed ed.). Urban & Fischer.
- Trail F. (2007). "Fungal cannons: explosive spore discharge in the Ascomycota". FEMS Microbiology Letterrs 276: 12–8.
- Pringle A, Patek SN, Fischer M, Stolze J, Money NP. (2005). "The captured launch of a ballistospore". Mycologia 97: 866–71. doi:10.3852/mycologia.97.4.866.
- Furlaneto MC, Pizzirani-Kleiner AA. (1992). "Intraspecific hybridisation of Trichoderma pseudokoningii by anastomosis and by protoplast fusion". FEMS Microbiology Letters 69: 191–5.
- Schardl CL, Craven KD. (2003). "Interspecific hybridization in plant-associated fungi and oomycetes: a review". Molecular Ecology 12: 2861–73.
- Bruns T. (2006). "Evolutionary biology: a kingdom revised". Nature 443: 758–61. doi:10.1038/443758a<a (inactive 2008-06-24).
- Lomako J, Lomako WM, Whelan WJ. (2004). "Glycogenin: the primer for mammalian and yeast glycogen synthesis". Biochim Biophys Acta. 1673: 45–55.
- Bowman SM, Free SJ. (2006). "The structure and synthesis of the fungal cell wall". Bioessays 28: 799–808.
- Mihail JD, Bruhn JN. (2005). "Foraging behaviour of Armillaria rhizomorph systems". Mycological Research 109: 1195–207.
- Shoji JY, Arioka M, Kitamoto K. (2006). "Possible involvement of pleiomorphic vacuolar networks in nutrient recycling in filamentous fungi". Autophagy 2: 226–7.
- Wu S, Schalk M, Clark A, Miles RB, Coates R, Chappell J. (2007). "Redirection of cytosolic or plastidic isoprenoid precursors elevates terpene production in plants". Nature Biotechnology 24: 1441–7.
- Xu H, Andi B, Qian J, West AH, Cook PF. (2006). "The α-aminoadipate pathway for lysine biosynthesis in fungi". Cellular Biochemistry and Biophysics 46: 43–64.
- Tudzynski B. (2005). "Gibberellin biosynthesis in fungi: genes, enzymes, evolution, and impact on biotechnology". Applied Microbiology and Biotechnology 66: 597–611.
- Siewers V, Smedsgaard J, Tudzynski P. (2004). "The P450 monooxygenase BcABA1 is essential for abscisic acid biosynthesis in Botrytis cinerea". Applied and Environmental Microbiology 70: 3868–76.
- Butterfield NJ. (2005). "Probable Proterozoic fungi". Paleobiology 31 (1): 165–82.
- Brundrett MC. (2002). "Coevolution of roots and mycorrhizas of land plants". New Phytologist 154 (2): 275–304.
- James TY et al (2006). "Reconstructing the early evolution of Fungi using a six-gene phylogeny". Nature 443: 818–22.
- Blackwell M, Vilgalys R, James TY, Taylor JW. (2008). Fungi. Eumycota: mushrooms, sac fungi, yeast, molds, rusts, smuts, etc. Version 21 February 2008.
- Eshet Y, Rampino MR, Visscher H. (1995). "Fungal event and palynological record of ecological crisis and recovery across the Permian-Triassic boundary". Geology 23 (1): 967-70.
- Foster CB.; Stephenson MH Marshall C, Logan GA, Greenwood PF. (2002). "A revision Of reduviasporonites Wilson 1962: description, Iillustration, comparison and biological affinities". Palynology 26 (1): 35–58.
- López-Gómez J, Taylor EL. (2005). "Permian-Triassic transition in Spain: a multidisciplinary approach". Palaeogeography, Palaeoclimatology, Palaeoecology 229 (1-2): 1–2.
- Looy CV.; Twitchett RJ, Dilcher DL, Van Konijnenburg-van Cittert JHA, Visscher H. (2005). "Life in the end-Permian dead zone". Proceedings of the National Academy of Sciences 162 (4): 653–9.
- Ward PD, Botha J, Buick R, De Kock MO, Erwin DH, Garrison GH, Kirschvink JL & Smith R (2005). "Abrupt and gradual extinction among late permian land vertebrates in the Karoo Basin, South Africa". Science 307 (5710): 709–14.
- White MM, James TY, O'Donnell K, Cafaro MJ, Tanabe Y, Sugiyama J. (2006). "Phylogeny of the Zygomycota based on nuclear ribosomal sequence data". Mycologia 98: 872–84.
- Remy W, Taylor TN, Hass H, Kerp H. (1994). "4-hundred million year old vesicular-arbuscular mycorrhizae". Proceedings of the National Academy of Sciences 91: 11841–3.
- Esser, K; Lemke PA (1994). The Mycota: A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research. Springer.
Lindahl BD, Ihrmark K, Boberg J, Trumbore SE, Högberg P, Stenlid J, Finlay RD. (2007). "Spatial separation of litter decomposition and mycorrhizal nitrogen uptake in a boreal forest". New Phytologist 173: 611–20. - Barea JM, Pozo MJ, Azcón R, Azcón-Aguilar C. (2005). "Microbial co-operation in the rhizosphere". Journal of Experimental Botany 56: 1761–78.
- Aanen DK. (2006). "As you reap, so shall you sow: coupling of harvesting and inoculating stabilizes the mutualism between termites and fungi". Biology Letters 2: 209–12.
- Nikoh N, Fukatsu T. (2000). "Interkingdom host jumping underground: phylogenetic analysis of entomoparasitic fungi of the genus Cordyceps". Molecular Biology and Evolution 17: 2629–38.
- Perotto S, Bonfante P. (1997). "Bacterial associations with mycorrhizal fungi: close and distant friends in the rhizosphere". Trends in Microbiology 5: 496–501.
- Arnold AE, Mejía LC, Kyllo D, Rojas EI, Maynard Z, Robbins N, Herre EA. (2003). "Fungal endophytes limit pathogen damage in a tropical tree". Proceedings of the National Academy of Science USA 100: 15649–54.
- Paszkowski U. (2006). "Mutualism and parasitism: the yin and yang of plant symbioses". Current Opinion in Plant Biology 9: 364–70.
- Hube B. (2004). "From commensal to pathogen: stage- and tissue-specific gene expression of Candida albicans". Current Opinion in Microbiology 7: 336–41. doi:10.1016/j.mib.2004.06.003.
- van der Heijden MG, Streitwolf-Engel R, Riedl R, Siegrist S, Neudecker A, Ineichen K, Boller T, Wiemken A, Sanders IR. (2006). "The mycorrhizal contribution to plant productivity, plant nutrition and soil structure in experimental grassland". New Phytologist 172: 739–52.
- Selosse MA, Richard F, He X, Simard SW. (2006). "Mycorrhizal networks: des liaisons dangereuses?". Trends in Ecology and Evolution 21: 621–8.
- Brodo, IM.; Sharnoff SD (2001). Lichens of North America. Yale University Press.
- Bisby GR, Ainsworth GC, Kirk PM, Aptroot A. (2001). Ainsworth & Bisby's Dictionary of the Fungi / by P. M. Kirk... [et al.]; with the assistance of A. Aptroot... [et al.]. Oxon: CAB International. p. 285.
- Jim Deacon (2005). Fungal Biology. Cambridge, MA: Blackwell Publishers. pp. 267–276.
- Jouquet P, Tavernier V, Abbadie L, Lepage M. (2005). "Nests of subterranean fungus-growing termites (Isoptera, Macrotermitinae) as nutrient patches for grasses in savannah ecosystems". African Journal of Ecology. 43: 191–6.
- Talbot NJ. (2003). "On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea". Annual Reviews in Microbiology 57: 177–202.
- Paoletti M, Buck KW, Brasier CM. (2006). "Selective acquisition of novel mating type and vegetative incompatibility genes via interspecies gene transfer in the globally invading eukaryote Ophiostoma novo-ulmi". Molecular Ecology 15: 249–62.
- Gryzenhout M, Wingfield BD, Wingfield MJ. (2006). "New taxonomic concepts for the important forest pathogen Cryphonectria parasitica and related fungi". FEMS Microbiology Letters 258: 161–72.
- Nielsen K, Heitman J. (2007). "Sex and virulence of human pathogenic fungi". Advances in Genetics 57: 143–73.
- Brakhage AA (2005). "Systemic fungal infections caused by Aspergillus species: epidemiology, infection process and virulence determinants". Current Drug Targets 6: 875–86.
- Kauffman CA. (2007). "Histoplasmosis: a clinical and laboratory update". Clinical Microbiology Reviews 20: 115–32.
- Cushion MT, Smulian AG, Slaven BE, Sesterhenn T, Arnold J, Staben C, Porollo A, Adamczak R, Meller J. (2007). "Transcriptome of Pneumocystis carinii during fulminate infection: carbohydrate metabolism and the concept of a compatible parasite".
- Kushwaha RKS, Guarro J. (eds). (2000). Biology of Dermatophytes and other ?Keratinophilic Fungi. Bilbao: Revista Iberoamericana Micologia. p. 177.
- Simon-Nobbe B, Denk U, Pöll V, Rid R, Breitenbach M. (2008). "The spectrum of fungal allergy". International Archives of Allergy and Immunology 145 (1): 58–86.
- Demain AL. (1991). "Production of beta-lactam antibiotics and its regulation". Proceedings of the National Science Council Republic of China Part B. 15: 251–65.
- Kulp, K. (2000). Handbook of Cereal Science and Technology. CRC Press.
- Piskur J, Rozpedowska E, Polakova S, Merico A, Compagno C. (2006). "How did Saccharomyces evolve to become a good brewer?". Trends in Genetics 22: 183–6.
- Kitamoto N, Yoshino S, Ohmiya K, Tsukagoshi N. (1999). "Sequence analysis, overexpression, and antisense inhibition of a β-xylosidase gene, xylA, from Aspergillus oryzae KBN616". Applied and Environmental Microbiology 65 (1): 20–4.
- "Trichoderma spp., including T. harzianum, T. viride, T. koningii, T. hamatum and other spp. Deuteromycetes, Moniliales (asexual classification system)". Biological Control: A Guide to Natural Enemies in North America.
- Joseph B, Ramteke PW, Thomas G. (2008). "Cold active microbial lipases: some hot issues and recent developments". Biotechnology Advances 26 (5): 457–70.
- Olempska-Beer ZS, Merker RI, Ditto MD, DiNovi MJ. (2006). "Food-processing enzymes from recombinant microorganisms—a review". Regulatory Toxicology and Pharmacology 45 (2): 144–58.
- Kumar R, Singh S, Singh OV. (2008). "Bioconversion of lignocellulosic biomass: biochemical and molecular perspectives". Journal of Industrial Microbiology and Biotechnology 35 (5): 377–91.
- Polizeli ML, Rizzatti AC, Monti R, Terenzi HF, Jorge JA, Amorim DS. (2005). "Xylanases from fungi: properties and industrial applications". Applied Microbiology and Biotechnology 67 (5): 577–91.
- van Egmond HP, Schothorst RC, Jonker MA. (2007). "Regulations relating to mycotoxins in food: perspectives in a global and European context". Analytical and Bioanalytical Chemistry 389: 147–57.
- Keller NP, Turner G, Bennett JW. (2005). "Fungal secondary metabolism - from biochemistry to genomics". Nature Reviews in Microbiology 3: 937–47. doi:10.1038/nrmicro1286.
- Demain AL, Fang A. (2000). "The natural functions of secondary metabolites". Advances in Biochemical Engineering/Biotechnology 69: 1–39.
- Rohlfs M, Albert M, Keller NP, Kempken F. (2007). "Secondary chemicals protect mould from fungivory". Biology Letters 3: 523–5
- Erdogan A, Gurses M, Sert S. (2004). "Isolation of moulds capable of producing mycotoxins from blue mouldy Tulum cheeses produced in Turkey". International Journal of Food Microbiology 85: 83–5.
- Leathem AM, Dorran TJ. (2007). "Poisoning due to raw Gyromitra esculenta (false morels) west of the Rockies". Canadian Journal of Emergency Medicine 9: 127–30.
- Karlson-Stiber C, Persson H. (2003). "Cytotoxic fungi—an overview". Toxicon 42: 339–49.
- Mythology and Folklore of Fly Agaric Paul Kendall, Trees for Life.
- López-Gómez J, Molina-Meyer M. (2006). "The competitive exclusion principle versus biodiversity through competitive segregation and further adaptation to spatial heterogeneities". Theoretical Population Biology 69: 94–109.
- Becker H. (1998). "Setting the Stage To Screen Biocontrol Fungi". United States Department of Agriculture, Agricultural Research Service.
- Keiller TS. "WHEY-BASED FUNGAL MICROFACTORY TECHNOLOGY FOR ENHANCED BIOLOGICAL PEST MANAGEMENT USING FUNGI"
- Deshpande MV. (1999). "Mycopesticide production by fermentation: potential and challenges". Critical Reviews in Microbiology 25: 229–43.
- Thomas MB, Read AF. (2007). "Can fungal biopesticides control malaria?". Nature Reviews in Microbiology 5: 377–83.
- Bush LP, Wilkinson HH, Schardl CL. (1997). "Bioprotective alkaloids of grass-fungal endophyte symbioses". Plant Physiology 114: 1–7.
- Bouton JH, Latch GCM, Hill NS, Hoveland CS, McCannc MA, Watson RH, Parish JA, Hawkins LL, Thompson FN. (2002). "Use of nonergot alkaloid-producing endophytes for alleviating tall fescue toxicosis in sheep". Agronomy Journal 94: 567–74.
- Douglas, M.S. (1995). Bioremediation of Contaminated Soils Using the White Rot Fungus Phanerochaete Chrysosporium.
- BBC. (2008). Fungi to fight 'toxic war zones'
- Fomina M, Charnock JM, Hillier S, Alvarez R, Livens F, Gadd GM. (2008). "Role of fungi in the biogeochemical fate of depleted uranium". Current Biology 18 (9): R375–7.
- Fomina M, Charnock JM, Hillier S, Alvarez R, Gadd GM. (2007). "Fungal transformations of uranium oxides". Environmental Microbiology 9 (7): 1696–710.
Further reading
- Alexopoulos CJ, Mims CW, Blackwell M, et al., Introductory Mycology, 4th ed. (John Wiley and Sons, Hoboken NJ, 2004)
- Arora D. (1986). Mushrooms Demystified: A Comprehensive Guide to the Fleshy Fungi. 2nd ed. Ten Speed Press.
- Deacon JW. (2005). Fungal Biology (4th ed). Malden, MA: Blackwell Publishers.
- Kaminstein D. (2002). Mushroom poisoning
Support our developers




