Mechanism of genetic recombination
Precocity theory of CD. Darlington is ordinarily discussed in context with meiotic chromosome pairing. However, Darlington extended this theory to explain recombination also. The theory assumes that prophase is precocious in meiotic cell division and therefore involves homologous pairing to satisfy the pairing need, which is achieved in mitosis due to duplication of chromosomes. This theory is now untenable in the light of recent information that DNA synthesis really takes place before the onset of prophase I in meiosis, although synthesis of a very small fraction of DNA called zygotene DNA (zyg DNA) (0.1 to 0.2%) is delayed till zygotene. Precocity theory presumes that DNA synthesis or chromosome duplication takes place later in pachytene or diplotene and then results in separation of homologous chromosomes (terminalization). On the basis of precocity theory, Darlington explained crossing over to be the result of strain or torsion produced due to coiling of homologous chromosomes and sister chromatids.
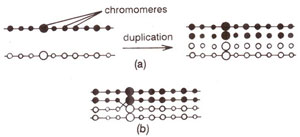
Fig. 10.4. Belling's model for crossing over showing duplication of chromomeres (a) followed by joining of chromomeres newly synthesized on two different homologous chromosomes (b).
In making a distinction between chiasmata and crossing over, we concluded earlier that crossing over or exchange of chromosome segments (which leads to genetic recombination) is the result of a breakage and reunion process. One may ask the question, whether it is actually a breakage-reunion process or only appear to be so, as envisaged in a hypothesis proposed by John Belling in 1928. While studying meiosis in some plant species, Belling visualized genes as beads (described as chromomeres), connected by nongenic inter-chromomeric regions.
Fig. 10.4. Belling's model for crossing over showing duplication of chromomeres (a) followed by joining of chromomeres newly synthesized on two different homologous chromosomes (b).
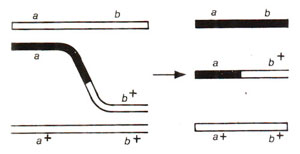
Fig. 10.5. Mechanism of genetic recombination based on copy choice mechanism.
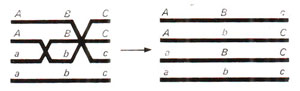
Fig. 10.6. A possible three strands double crossover, making Belling’s copy-choice (switch) model very unlikely.
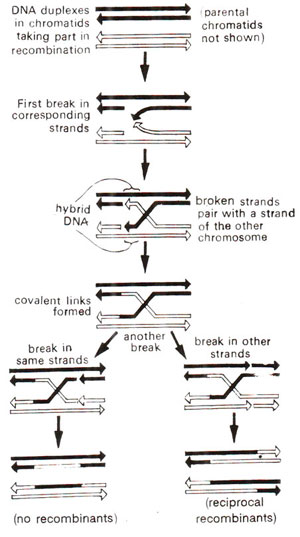
Fig. 10.7. Mechanism of recombination as explained on the basis of hybrid DNA model of R. Holliday (1964).
Fig. 10.5. Mechanism of genetic recombination based on copy choice mechanism.
Fig. 10.6. A possible three strands double crossover, making Belling’s copy-choice (switch) model very unlikely.
Fig. 10.7. Mechanism of recombination as explained on the basis of hybrid DNA model of R. Holliday (1964).
During 1960s, hybrid DNA models had become-very popular, since these models best explained the results obtained in certain microbial systems. In these models, only one strand in each of two DNA duplexes belonging to non-sister chromatids (from homologues) breaks. The single strands released from these breaks then pair crosswise with unbroken strands by complementary base pairing. This results in the formation of hybrid DNA segments, and hence the name hybrid DNA models.
Several hybrid DNA models have been proposed during the last three decades. Two of the earliest models included the one proposed in 1963 by H.L.K. Whitehouse of Cambridge, England and the other in 1964 by R. Holliday of London. The two models mainly differ in so far as Whitehouse believed that breaks would occur in single strands having opposite polarity, while Holliday proposed that breaks would occur in sirands having same polarity. The details of Holliday's model, which is relatively simple and has been widely accepted, are presented in Figure 10.7. In contrast to these models involving single strand breaks, there are also hybrid DNA models assuming double strand breaks, which are discussed in Molecular Mechanisms of Recombination.
Support our developers




