Detection of Mutations in Drosophila
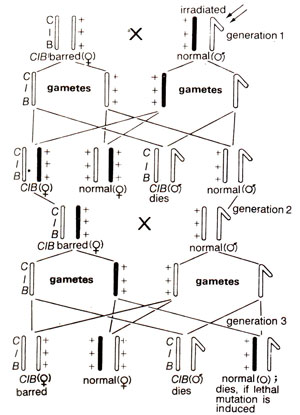
Fig. 21.3. Muller's CIB method for detection of sex linked lethal mutations in Drosophila.
The ClB method. This method involves use of a ClB stock which carries (i) an inversion in heterozygous state to work as crossover suppressor (C), (ii) a recessive lethal (l) on X-chromosome in heterozygous state, and (iii) a dominant marker, Barred (B)for the barred eye (narrow eye). One of the two X-chromosomes in a female fly carried all these three features and the other X-chromosome was normal. Male flies irradiated for induction of mutations were crossed to ClB females. Male progeny receiving ClB X-chromosome will die (Fig. 21.3).
Fig. 21.3. Muller's CIB method for detection of sex linked lethal mutations in Drosophila.
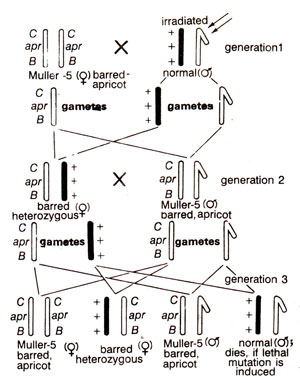
Fig. 21.4. Muller-5 method for detection of sex linked mutations in Drosophila.
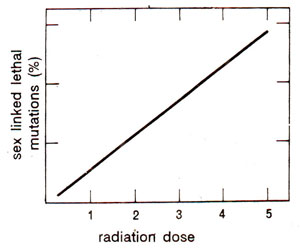
Fig. 21.5. A linear relationship between the frequency of sex linked lethal mutations and the dose of radiation in kilo Roentgens (kR)
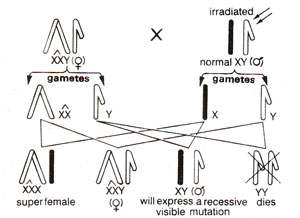
Fig. 21.6. Attached X-method for detection of sex linked visible mutations.
Fig. 21.4. Muller-5 method for detection of sex linked mutations in Drosophila.
Fig. 21.5. A linear relationship between the frequency of sex linked lethal mutations and the dose of radiation in kilo Roentgens (kR)
Detection of sex linked visible mutations Attached X-method. The methods described in the preceding section were meant for detection of sex linked lethals. For detection of sex linked visible mutations, Muller-5 and attached X-chromosomes were used. The attached X females (XXY) have a special advantage. When these females are crossed to an irradiated male, X-chromosome of irradiated male goes either to superfemale daughters or to the sons. Since in sons there is a single X-chromosome, any visible induced mutation will immediately express itself and can be easily scored (Fig. 21.6).
Fig. 21.6. Attached X-method for detection of sex linked visible mutations.
Balanced lethal system and detection of mutations. Detection of mutations on autosomes (chromosomes other than sex chromosomes) makes use of a balanced lethal stock. Balanced lethal stock is one which carries recessive lethal alleles on two specific homologous chromosomes, their normal allele being present on its homologue; consequently homozygous individuals will not survive. The concept of balanced lethal would be better understood using a specific example. For detection of visible mutations in Drosophila, a stock carried the dominant genes Cy (curly wing)and L (lobed eye)on one chromosome and Pm (plum eye i.e. brownish eye) on the other homologue so that the organism can be designated as Cy L/Pm. Phenotype of the stock which was heterozygous for Cy, L and Pm, was curly, lobe and plum. If such individuals are crossed among themselves, progeny is always heterozygous, because each of the two homologous chromosomes carried recessive lethal genes (Fig. 21.7). Through the presence of inversion, crossing over was also suppressed, so that Cy and/or L may not be transferred to the chromosome carrying Pm and vice versa.
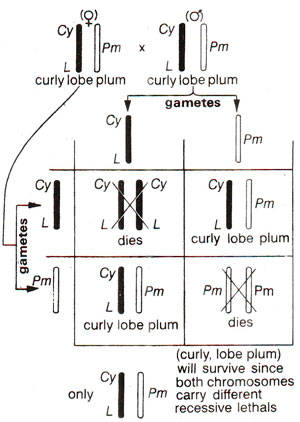
Fig. 21.7. Balanced lethal system in Cy LIPm Drosophila.
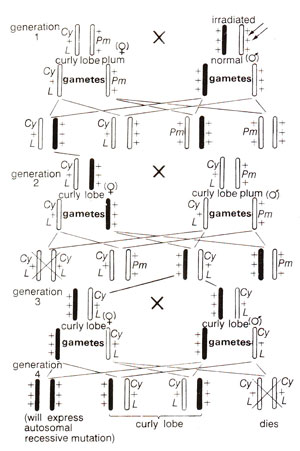
Fig. 21.8. Detection of autosomal mutations in Drosophila.
Support our developers




