Polyploidy
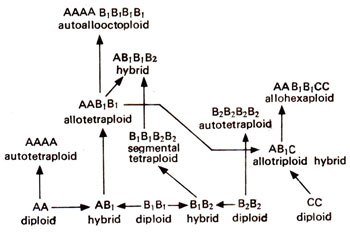
Fig. 20.8. Different kinds of polyploids and their derivations.
Fig. 20.8. Different kinds of polyploids and their derivations.
(a) Origin and production of autopolyploids.
The autopolyploids may occur in nature or may be artificially produced. When they are found in nature, their autopolyploid nature is inferred mainly by their meiotic behaviour. One of the very common examples of natural autopolyploidy relevant to Northern India, pertains to common 'doob' grass (Cynodon dactylon). In U.P. and Bihar, common 'doob' grass was found to be an autotriploid as inferred from its meiotic behaviour (Gupta and Srivastava, 1970).It is perhaps successful due to efficient vegetative reproduction, because, as will be seen, autoeupolyploids are normally triploids and set no seeds. Autotriploids are also known in watermelons, sugarbeet, tomato, grapes and banana, although in several of these cases the polyploids have been artificially produced. Similarly autotetraploids are known in rye (Secale cereale), corn (Zea mays), red clover (Trifolium pratense), berseem (Trifolium alexandrium), marigolds (Tagetes), snapdragons (Antirrhinum), Phlox, grapes, apples, etc. In Oenothera lamarckiana, an American plant, a giant mutant described by Hugo de Vries was later discovered to be an autotetraploid.
In many cases listed above, autopolyploidy is artificially induced. Since polyploids are normally larger and more vigorous, their role in crop improvement has been realized and techniques developed for artificial induction of polyploidy. Polyploidy is mainly induced by treatment with aqueous solution of a drug called colchicine. This drug has the property of arresting and breaking the spindle so that a cell division without cell wall formation may be affected leading to doubling of chromosome number. The concentration of aqueous solution of colchicine may vary from 0.01 % to 0.50% and the treatment may be given in one of the following manners, (i) Seed treatment may be mainly given by soaking seeds for different durations in. aqueous solution of colchicine. (ii) Injections of colchicine solution may also be given at seeding stage so as to inject solution into cortex tissue with the help of a hypodermic needle, (iii) Axiliary bud treatment is also effective. Since bud is meristematically active, placing cotton soaked in colchicine on the bud and continuous dropping of solution on the bud leads to induction of polyploidy in the branch arising from the treated bud. (iv) Shoot apex treatment is brought about just like bud treatment and is fairly effective, but the shoot apex should come in direct contact of the solution. In order to facilitate this, young leaves covering the shoot apex may be removed.
One of the important effects of polyploidy is to produce 'gigantism'. The autopolyploids may be normally larger in size. Sometimes plants may be smaller than diploids, but leaves, flowers and the cells themselves may be bigger in size. Some of the important effects of polyploidy are a's follows : (i) With increase in cell size, water content increases leading to decrease in osmotic pressure. This results into loss of resistance against frost, etc. (ii) Growth rate decreases due to slower rate of cell division; this leads to a decrease in auxin supply and a decrease in respiration, (iii) In autopolyploids, time of blooming is delayed and prolonged due to slow growth rate, (iv) At higher ploidy level (autooctoploids or higher), the adverse effects are highly pronounced and lead to death of plants.
(d) Cytology of autopolyploids.
In an autopolyploid, there will be more than two sets of homologous chromosomes. This leads to formation of multivalents instead of bivalents as found in diploids. An important difference exists even between autotriploids and autotetraploids, because while in the latter normal disjunction is possible giving rise to diploid gametes, in triploids it is not possible. In an autotriploid, there are three sets of homologous chromosomes. If these three sets are normally paired, trivalents can not disjoin normally and will either disjoin 2 : 1 chromosomes to two poles or will disjoin 1 : 1 leaving one chromosome as a laggard. The number of chromosomes in gametes of triploid organism, therefore, will vary from n to 2n. Most of these gametes are unbalanced leading to high degree of sterility.
(e) Genetics of autopolyploids (trisomic and tetrasomic inheritance).
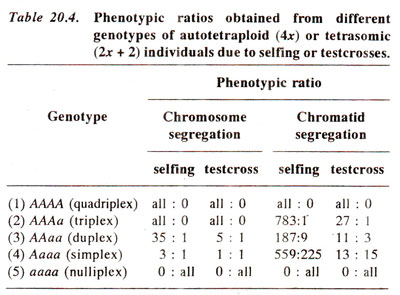
The segregation pattern in polyploids is quite different than what we find in diploids. In a polyploid there are more than one kind of heterozygotes, because there are more than two homologous chromosomes. The different possible genotypes and expected ratios in autotetraploids are given in Table 20.4. Similar ratios for triplqids are given in Table 20.1. The ratios are based on the assumption that either the gene is close to centromere leading to what we call chromosome segregation or the gene is away from centromere so that crossing over between gene and centromere will take place and ratios will be modified. In the latter case it will be called chromatid segregation, because segregation is taking place not at chromosome level, but at chromatid level due to crossing over.
(f) Uses of induced polyploidy
Since fertility level and seed set are low in induced polyploids, their utility in improvement of crop plants will depend on two major considerations, namely : (i) what is the commercial product, because if seed is not the commercial product low seed set will not be a handicap, and (ii) whether or not fertility level is important. Keeping these requirements in mind, seedless fruits can be produced by using triploids as in case of seedless watermelons produced by H. Kihara (who died in 1986) in Japan. These triploids were obtained from seeds raised by a cross tetraploid x diploid, the tetraploids being raised from diploids by colchicine treatment. This method was also used for obtaining triploids in sugarbeet, tomato and grapes.
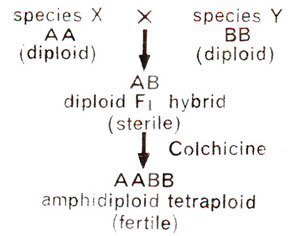
Fig. 20.9. Derivation of a tetraploid amphidiploid from two diploid species.
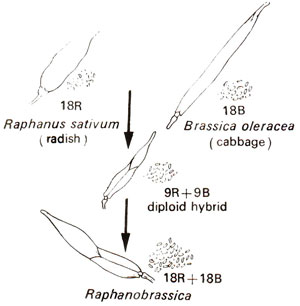
Fig. 20.10. Artificial synthesis of Raphanobrassica.
Polyploidy may also result from doubling of chromosome number in a F1 hybrid which is derived from two distinctly different species. This will bring two different sets of chromosomes in F1 hybrid. The number of chromosomes in each of these two sets may differ. Let A represent a set of chromosomes (genome) in species X, and let B represent another genome in a species Y. The F1 will then have one A genome and another B genome. The doubling of chromosomes in this F1 hybrid (AB) will give rise to a tetraploid with two A and two B genomes (Fig. 20.9). Such a polyploid is called an allopolyploid or amphidiploid.
Fig. 20.9. Derivation of a tetraploid amphidiploid from two diploid species.
Raphanobrassica is a classical example of allopolyploidy. In 1927, G.D. Karpechenko, a Russian geneticist, reported a cross between Raphanus sativus (2n = 18) and Brassica oleracea (2n = 8) to produce F1hybrid which was completely sterile. This sterility was due to lack of chromosome pairing, since there is no homology between genomes from Raphanus sativus and Brassica oleracea. Among these sterile F1 hybrids, Karpechenko found certain fertile plants. On cytological examination these fertile plants were found to have 2n = 36 chromosomes, which showed normal pairing into 18 bivalents (Fig. 20.10).
Fig. 20.10. Artificial synthesis of Raphanobrassica.
Common cultivated wheat is another important example of allopolyploidy, although its allopolyploid nature has now been questioned. There are three different chromosome numbers in the genus Triticum, namely 2n = 14, 2n = 28 and 2n - 42. The common wheat is hexaploid with 2n = 42, and is derived from three diploid species : (i) AA = Triticum aegilopoides (2n = 14), (ii) BB = Aegilops speltoides ? (2n = 14) (in the past evidence was made available, showing that Ae. speltoides may not be the donor of B genome; it is also believed that the donor of B genome may never be discovered) and (iii) DD = Aegilops squarrosq (2n = 14). The hexaploid wheat, therefore, is designated as AABBDD, the tetraploid (2n = 28) as AABB and diploid (2n = 14) as AA. There is, however, evidence available now which suggests that A, B and D genomes from three diploid species mentioned above are not much different from one another, so that it is believed that the three diploid progenitors of common hexaploid wheat were derived from a common ancestor. For this reason, the common hexaploid wheat is now considered an autopolyploid rather than an allopolyploid. At best, it may be a segmental allopolyploid.
Certain allopolyploids were artificially produced in order to find out the origin of naturally occurring allopolyploids. Common hexaploid wheat and tetraploid cotton furnish two such examples. These two and few other examples will be briefly discussed.
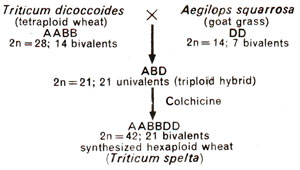
Fig. 20.11. Artificial synthesis of hexaploid wheat.
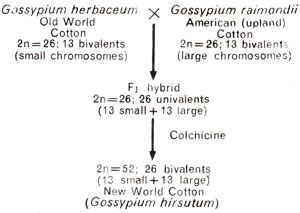
Fig. 20.12. Artificial synthesis of New World cotton.
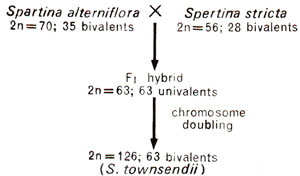
Fig. 20.13. Probable origin of Spartina townsendii.
Fig. 20.11. Artificial synthesis of hexaploid wheat.
(2) Gossypium hirsutum, popularly known as upland cotton, is another interesting example of amphidiploidy. Old World cotton has 13 pairs of large chromosomes, while American or upland cotton has 13 pairs of smaller chromosomes. The New World cotton, the cultivated long-staple type has 26 paris of chromosomes, 13 large and 13 small. J.O. Beasley crossed the American and Old World cottons and doubled the chromosome number in the F1 hybrids (Fig. 20.12). The amphidiploid thus produced resembled the cultivated New World cotton and when crossed with it gave fertile F1 hybrids. The evidence presented in Figure 20.12, thus showed that New World cotton, the cultivated Gossypium hirsutum (tetraploid) originated from two diploid species, namely G. herbaceum (2n = 26) and G. raimondii (2n = 26).
Fig. 20.12. Artificial synthesis of New World cotton.
Fig. 20.13. Probable origin of Spartina townsendii.
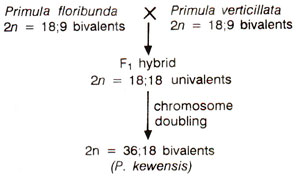
Fig. 20.14. Probable origin of Primula kewensis.
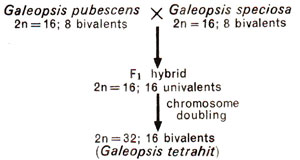
Fig. 20.15. Probable origin of tetraploid of Galeopsis tetrahit.
Fig. 20.14. Probable origin of Primula kewensis.
(5) Galeopsis tetrahit (2n = 32) could be synthesized (Fig. 20.15) from crosses involving diploid species G. pubescens (2n = 16) and G. speciosa (2n = 16). However, A. Muntzing, who performed these experiments suggested that G. tetrahit is not a strict amphidiploid, but has a tendency towards autopolyploidy.
Fig. 20.15. Probable origin of tetraploid of Galeopsis tetrahit.
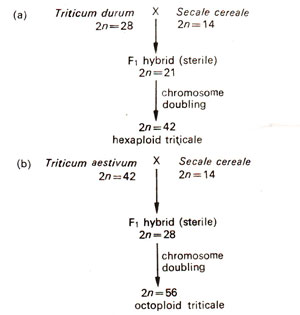
Fig. 20.16. Artificial synthesis of (a) hexaploid triticale and (b) octoploid triticale.
In recent years, considerable emphasis has been laid on the possibility of utilizing a new man made cereal known as triticale, on a commercial scale. It is already grown in an estimated area of more than one million hectares and research at several places all over the world is in progress to improve this man made crop. Work done in this crop has been reviewed in detail (Muntzing, 1979, Gupta and Priyadarshan, 1982; Gupta and Reddy, 1991). Several hundred cultivars of triticale have been released during the last 10 years and more cultivars of this crop are being released every year. The area under its cultivation is also fast increasing (for more details consult, Gupta, 1984, 1985, 1987; Gupta and Reddy, 1991).
Fig. 20.16. Artificial synthesis of (a) hexaploid triticale and (b) octoploid triticale.
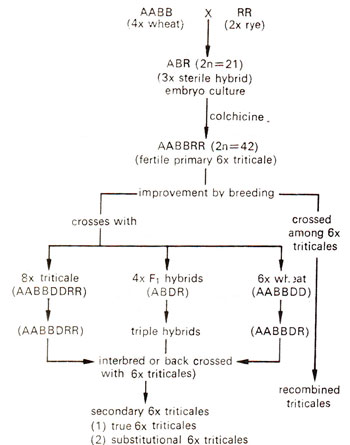
Fig. 20.17. Methods of improvement of primary triticales to obtain secondary and recombined triticales (from Gupta, 1984).
Fig. 20.17. Methods of improvement of primary triticales to obtain secondary and recombined triticales (from Gupta, 1984).

A more detailed iist of more than 150 triticales was published by CIMMYT (International Centre for Wheat and Maize Improvement) in 1986 (Spring Triticale, Names, Pedigrees, Origins : compiled by O.S. Abdalla et al.)
In some allopolyploids, the different genomes which are present are not quite different from one another. Consequently, in these polyploids chromosomes from different genomes do pair together to some extent and multivalents are formed. This mearts that segments of chromosomes and not the whole chromosomes are homologous. Therefore, such allopolyploids are called segmenta! allopolyploids according to Stebbins (1943, 1950). These segmental allopolyploids are intermediate between autopolyploids and allopolyploids and can be identified by their meiotic behaviour.
It is also believed that most of the naturally occurring polyploids are neither true autopolyploids nor true allopolyploids but are rather segmental allopolyploids. Our common hexaploid bread wheat is also regarded to be a segmental ailopolyploid, because the three diploid genomes (A, B and D) are related (homoeologous) to each other.
Support our developers




