X/A ratio and multiple numerator elements (Drosophila and Coenorhabditis)
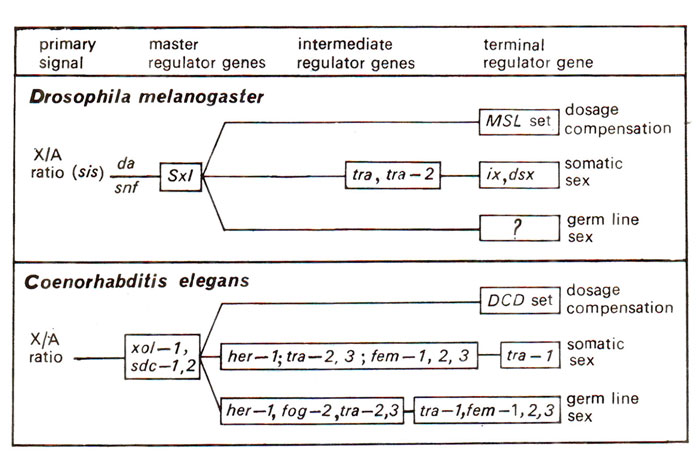
Fig. 17.10. General organization of sex determination and sex differentiation genes in Drosophila melanogaster and C. elegans.
Fig. 17.10. General organization of sex determination and sex differentiation genes in Drosophila melanogaster and C. elegans.
In the X/A ratio, X is the numerator and A is the denominator and there are genetic elements on both, described as 'numerator elements' and 'denominator elements' that contribute to this ratio. The elements interact with the target gene of X/A ratio signal. For instance,
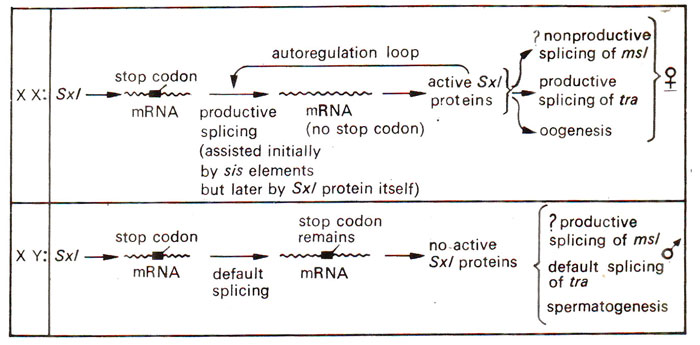
Fig. 17.11. Probable role of Sxl gene (and regulation of its activity) in sex determination and sex differentiation in Drosophila melanogasler.
(a) Sex linked master control genes. As discussed above, the X/A ratio works as a signal and sets the master control genes into functional state. These master genes include Sxl (sex lethal)gene in Drosophila and xol-1, sdc-1 and sdc-2genes in C. elegans. In Drosophila, Sxl responds to the dosage of sis-a and sis-b genes and also to other genes like da (daughterless) and snf (sans-fille).
Fig. 17.11. Probable role of Sxl gene (and regulation of its activity) in sex determination and sex differentiation in Drosophila melanogasler.
The function of Sxl is achieved by differential control on splicing of its transcript (HnRNA), which has a stop codon (in frame) undergoing one of the following two fates : (i) in males, due to stop codon, full length protein is not obtained, and the truncated proteins produced are non-functional; (ii) in females during early phase, stop codon is removed by splicing (due to the activation by sis elements),
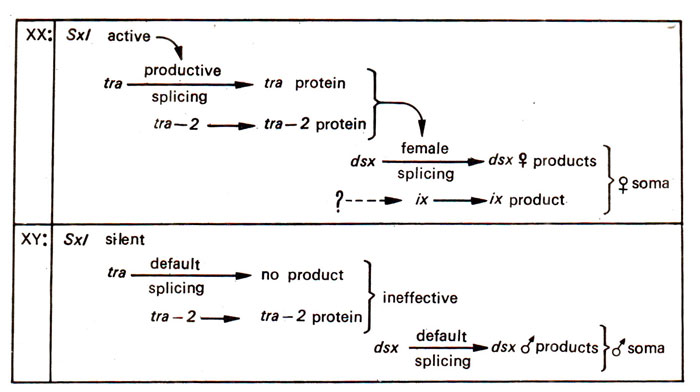
Fig. 17.12. Somatic cascade in Drosophila melanogaster; in XX flies, Sxl promotes productive splicing of tra transcript to produce tra protein which in association with tra-2 promotes female specific splicing of dsx transcripts and finally dsx product in association with ix product leads to female somatic development; in XY flies both tra and dsx adopt a default mode of splicing.
Fig. 17.12. Somatic cascade in Drosophila melanogaster; in XX flies, Sxl promotes productive splicing of tra transcript to produce tra protein which in association with tra-2 promotes female specific splicing of dsx transcripts and finally dsx product in association with ix product leads to female somatic development; in XY flies both tra and dsx adopt a default mode of splicing.
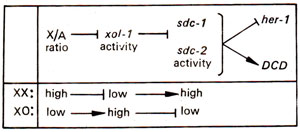
Fig. 17.13. The roles of XO lethal gene (xol-1)and sex and dosage compensation genes (sdc-1, sdc-2)in C. elegant;mutations in these two classes of genes have essentially opposite effects; sdc mutations are epistatic to xol-1 mutations (barred arrows i.e. —| indicate negative interaction or down regulation).
(a) Sex linked master control genes. In C. elegans, X/A ratio works as a signal for a master control gene having functional similarity to Sxl. This gene is sdc-2 (sex and dosage compensation) which is sex linked (located on X) and activates femaleness. This gene works with another gene sdc-1, so that both sdc-1and sdc-2are under negative control of xol-1(XO lethal) in XO animals (although xol-1mutations affect both XX and XO animals). This gene xol-1is compared with da (daughterless) in fruitfly, and its activity is negatively correlated with high or low X/A ratio (Fig. 17.13). The interactions in C. elegans are thus more complex than in fruitfly.
Fig. 17.13. The roles of XO lethal gene (xol-1)and sex and dosage compensation genes (sdc-1, sdc-2)in C. elegant;mutations in these two classes of genes have essentially opposite effects; sdc mutations are epistatic to xol-1 mutations (barred arrows i.e. —| indicate negative interaction or down regulation).
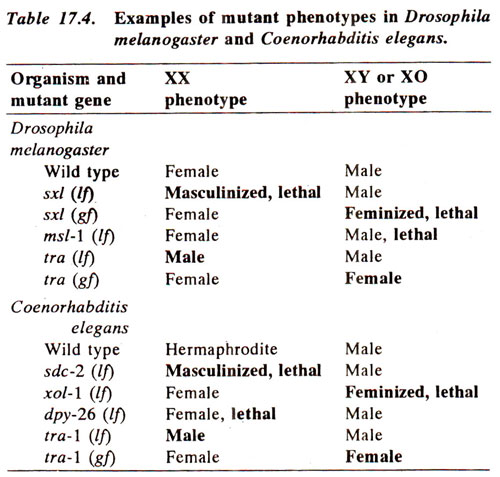
The same seven genes that control somatic sex in C. elegans, also control germ line sex, but following differences are noteworthy : (i) fern genes have dual role, masculinizing directly by inducing spermatogenesis and by down regulating tra-1(in the absence of fern gene activity, only oocytes are produced, irrespective of the state of tra-1); (ii) the role of tra-1is complicated in gametogenesis, but some level of tra-1activity is necessary for gametogenesis in both sexes; (iii) fog-2 (feminization of germ line)down regulates tra-2 transiently, making fern genes active, thus promoting spermatogenesis. The tra-2 activity is under general negative control in XO animals (presumably by her-1)and under germ line specific negative control in XX animals (presumably by fog-2). A model showing interactions involved in the germ line development are shown in Figure 17.15.
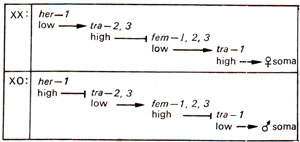
Fig. 17.14. Somatic cascade inferred for C. elegans; her (hermaphroditization gene), fern (feminization gene); tra wild allele feminizes; the seven genes involved adopt opposite activity states in the two sexes; in XX animals, her-1activity is low and in XO it is high, which ultimately ends up with high activity of tra-genes in XX leading to femaleness and low activity of tra genes in XO leading to maleness.
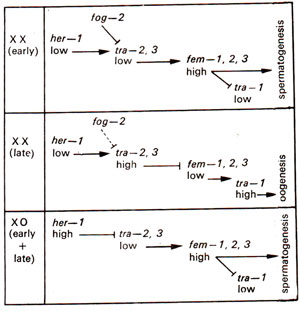
Fig. 17.15. Germ-line cascade in C. elegans; low her-1activity in XX leads to hermaphrodite, while high her-1activity in XO leads to male; high fern activity leads to spermatogenesis and low fern activity leads to oogenesis; mutants at fog-2and some tra-2mutants (gf = gain of function) eliminate spermatogenesis from XX rendering them female, but they do not affect XO individuals, so that XX female and XO male system develops.
Support our developers




