Simple Tissues and Complex Tissues
Another way whereby tissues can be classified is agian into two kinds: simple and complex. Simple tissue is composed of only one kind of cell, and complex tissue is composed of two or more kinds of cells.Simple Tissues
There are five types of simple tissue: epidermis, parenchyma, collenchyma, sclerenchyma, and cork Epidermis covers the outer surfaces of leaves, young roots, and stems. As plants (especially woody plants) age, the epidermis (a primary tissue) on stems and roots is replaced by bark (a secondary tissue). Stomates are found with epidermal cells on leaves and stems but not on roots. Epidermal cells are living. They are almost entirely covered on the outer surface by a waxy cuticle, which retards water passage.
 |
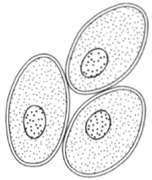
| Figure 26-2 Collenchyma |
 |
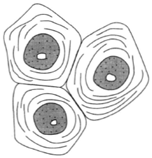
| Figure 26-1 Parenchyma, an unspecialized living tissue having conspicuous intercellular spaces. |
Parenchyma cells are commonly isodiametric (as broad as they are long) and have thin walls, living protoplasts, and conspicuous intercellular spaces. Parenchyma is regarded as an unspecialized tissue. If the cells have chloroplasts, the tissue is called chlorenchyma. In potatoes and fruits, the parenchyma cells store starch. Parenchyma is the main tissue of apples, pears, plums, and tomatoes. One of the most significant features of parenchyma is that it retains living protoplasts at maturity.
Collenchyma is the first of the supporting tissues found in young stems and leaves. These cells retain living protoplasts in maturity, as do parenchyma cells. Wall thickenings are conspicuous at the corners of cells. While these thickenings contribute to the strength of cells, they also remain plastic so the cells can stretch. The walls are composed primarily of cellulose but also contain some pectic compounds. These cells become elongated (often as many as two millimeters long) and frequently have chloroplasts. Intercellular spaces are sometimes noted.
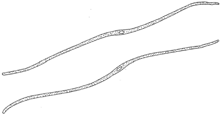 |
| Figure 26-3 Fibers. Note the small lumen in the center of the cell. |
There are two kinds of sclerenchyma, another supporting tissue: stone cells and fibers. Stone cells (also called sclereids) are short, nearly isodiametric, and have thick, lignified walls with tubular pits. They occur in small clusters in the tissue of pears. They also compose the hard shells of walnuts. In maturity, these cells do not have any living contents. In this sense, they can be distinguished from collenchyma.
Fibers are much elongated and have greatly thickened secondary walls, which may be of cellulose and lignin. The walls are often so thick that the lumen of the cell nearly disappears. There are no living protoplasts. Fibers are of two kinds: those that occur in wood and those that occur elsewhere. The latter are called bast fibers, and they are also of two kinds: those having lignified cell walls and those having cellulose cell walls. The latter are valued commercially. Bast fibers are often associated with the phloem, or inner bark,
 |
| Figure 26-4 Sclereids (stone cells) |
and may reach lengths of several feet-that is, an individual cell reaches a length of several feet. Examples include sisal, jute, hemp, ramie, hennekan, sansievaria, maguey, palma, formium, caroa, coir, latona, esparto, amuritius, and cabuya.
The last of the simple tissues is cork. Cork cells are found on the surfaces of stems and roots of the older parts of plants. Cork cells are flattened, and their walls are impregnated with the fatty substance suberin. There are no intercellular spaces, and the protoplasts soon die.
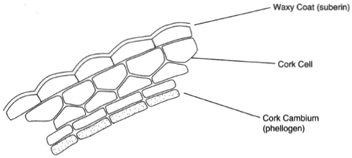 |
| Figure 26-5 cork |
Complex Tissues
There are two kinds of complex tissues: xylem and phloem. Both are conducting tissues, xylem being involved in the movement of water from the roots to the upper parts of the plant and phloem being involved in transporting the products of photosynthesis. These tissues are somewhat different in angiosperms than in gymnosperms.
Xylem and Phloem in Angiosperms. Several tissues are derived from the apical meristem; one of these is procambium, which gives rise to the cambium. In a woody plant, the cambium is a cylinder of meristematic cells one cell in thickness and lying just below the bark. Its divisions produce xylem toward the center and phloem toward the outside. The xylem has two kinds of conducting cells, tracheids and vessels, and two kinds of nonconducting cells, fibers and parenchyma.
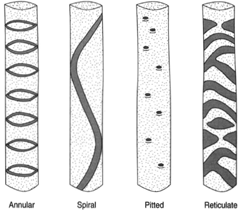 |
|
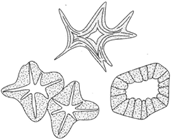
Figure 26-6 Types of tracheids. |
The tracheids, vessels, and fibers are nonliving when functional. kacheids are tapered at their ends and make contact with other tracheids. The secondary wall thickenings in tracheids produce a variety of patterns. Some forms show rings, some continuous spirals. In others, the wall thickenings produce a reticulate pattern, or a pitted appearance. A variety of names can therefore be applied to tracheids: annular tracheids, spiral tracheids, scalariform tracheids, reticulate tracheidsa, nd pitted tracheids (see figure 26-6).
Vessels are composed of vessel elements, individual cells with the end walls digested away. Vessels formed in this way can be up to one meter long. The ends of vessels have walls at top and bottom. These end walls are perforated vessels are thus able to conduct water more readily than are tracheids.
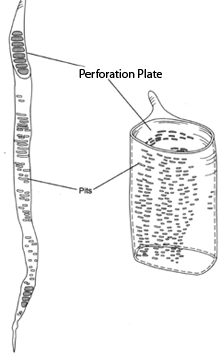 |
| Figure 26-7 Vessel elements. The evolutionary trend seems to be toward shorter and wider elements as at right. |
There are also fibers and parenchyma in xylem. The fibers are greatly elongated and have thick secondary walls such that the lumen is nearly obliterated. The parenchyma cells, the only living cells in xylem, are often arranged along radial lines, resulting in rays. They may be one to several cells wide and several to many cells high. These cellas re the products of cambial activity just as are the tracheids and vessels. As the circumference of a stem increases, new rays are produced. (Further details are provided in chapter 32 on stems.)
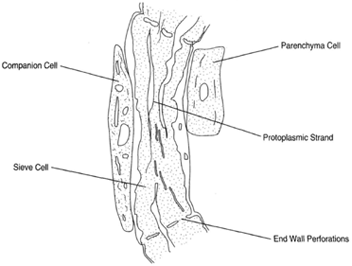
In angiosperms, the phloem (which is involved in 'transporting the products of photosynthesis from the leaves to the other parts of the plant) is distinguished by one particular feature: the sieve tubes. These are composed of elongated slender cells, the sieve-tube elements, having perforations in the end walls and positioned in a manner to produce continuous channels. Secondary wall thickenings do not occur, and the cells retain living cytoplasm although the nucleus is absent. Cytoplasm runs through the perforations in the cells to connect with adjacent sieve-tube elements. A central vacuole persists and extends through the end-wall perforations. The vacuolar membrane is not apparent.
 |
|
Figure 26-8
Sieve cell with companion cell.
The protoplasmic strand runs through the end-wall perforations. |
Another feature of angiosperm phloem is the companion cell, which lies beside each sieve-tube element. This is a parenchymatous, living cell. The sieve-tube element and companion cell are derived from the division of a parent cell, which can be called the sieve-tube mother cell. The cytoplasm of these two cells remains in contact by delicate strands that run through microholes in the walls. These are plasmodesmata. There may be a single com panion cell lying beside each sieve-tube element or several cells, which arise by subsequent divisions of the companion cell.
A complex carbohydrate composed of glucose units and called callose tends to be deposited at the site where cell perforations are most developed. This deposition eventually results in closing off the holes, thereby rendering the sieve tubes nonfunctional.
In addition to the sieve tubes and companion cells the phloem also contains fibers and ordinary parenchyma.
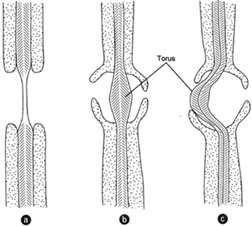 |
| Figure 26-9 (a) A pit as it occurs in angiosperms. (b) A bordered pit as seen in gymnosperms. (c) The torus, here deflected and appearing to close the pit mouth on one. |
The movement of water and its dissolved materials through the phloem is not thoroughly understood. The solutions can move in opposite directions at different times, and different solutes can move in opposite directions through the same tubes at the same time. This transport is achieved by living cells, and it is clear that energy is expended in the process. If phloem cells are killed, the movement ceases.
Xylem and Phloem in Gymnosperms. When the xylem of gymnosperms is compared to that of angiosperms, the most conspicuous difference is the of lack vessels. The only conducting cells in the wood of gymnosperms are the tracheids. It is yet possible to discern annual rings because the tracheids formed in the spring are wider in diameter than those produced later in the season.
The pits formed in the secondary walls of the tracheids are bordered pits quite unlike those of angiosperms (see figure 26-9). The primary wall in the center of the pit is thickened it is called a torus. The secondary wall produces borders. The pitso f adjacent cells are directly opposite, at the end pits are sometimes collectively called a pit pair. hIta s been observed that the torus can be deflected and press closely to the mouth of the pit, thereby obstructing entry into the pit cavity. This seems to be without significance, however.
The phloem of gymnosperms is different from that of angiosperms in that sieve tubes are not formed. Sieve cells occur, but the end walls are not perforated as they are in angiosperms. Furthermore, there are no companion cells associated with the sieve cells in gymnosperms.
Support our developers




