DNA Replication
DNA replication of the E. coli chromosome begins at a single origin of replication (oriC) and proceeds bidirectionally to a termination site located approximately halfway around the circular chromosome. During replication, the DNA strands of the double helix must be both unwound and separated.DNA replication is initiated when a protein encoded by the gene DNA A binds repetitive 9-mer sequences at the origin. Subsequently, helicases specified by DNA B and inhibitory proteins encoded by DNA C bind repetitive 13-mer sequences. As helicase progresses 5' to 3', dissociation of protein DNA C allows the helicase to unwind the DNA . The unwinding produces positive superhelical turns in the rest of the DNA , making it energetically favorable to continue unwinding the strands. To unwind the DNA , positive superhelical turns have to be removed by cutting the DNA and allowing it to relax or by introducing negative superhelical turns to compensate for the positive ones. The introduction of negative superhelical turns requires energy and an enzyme called DNA gyrase (a topoisomerase). DNA gyrase is an enzyme that can both remove positive supercoils or introduce negative supercoils into the DNA and thereby make strand separation energetically more favorable. Presumably the DNA gyrase binds ahead of the unwound DNA during replication. Single-stranded binding proteins (SSBPs) act to temporarily stabilize the unwound state.
DNA replication (Figure 3-2) begins with the synthesis of a 30 nucleotide long RNA primer by an RNA polymerase called primase (specified by DNA G). The helicase and primase subsequently form a complex enzyme system known as the primosome, which synthesizes primers after DNA synthesis begins. Two catalytic subunits of DNA polymerase III (PolC) associate with the templates and the 3' ends of the primers and begin to polymerize deoxyribonucleotides into DNA . DNA gyrase continues to remove positive supercoils and/or introduces negative supercoils ahead of the primosome that is opening the two strands of DNA . At various intervals, the template signals the primase portion of the primosome to polymerize primer RNAs about 30 nucleotides long on only one template at the replication fork. DNA polymerase III polymerizes DNA 5' to 3' from each of the primers at the replication fork. One strands of DNA is polymerized toward the replication fork and continues to be elongated as the DNA unwinds further. The second strand of DNA is polymerized away from the replication fork. As the DNA unwinds further, a new primer is synthesized away from the replication fork and the DNA polymerase synthesizes DNA from the last primer toward the previous RNA primer. As the DNA polymerase reads the template strand, it selects complementary nucleotides for the nascent strand based on hydrogen bonding capability.
The DNA synthesized toward the replication fork is synthesized in a continuous manner and is called the leading strand. The opposite DNA strand is synthesized in a discontinuous manner away from the replication fork and is referred to as the lagging strand. The leading and lagging strands are synthesized halfway around the bacterial chromosome until they encounter the lagging and leading strands synthesized at the other replication fork.
The RNA-DNA fragments that initially constitute the lagging strand are known as Okazaki fragments, named after the scientist who discovered them.
The RNA primers are removed by a DNA repair enzyme called DNA polymerase I specified by polA. It uses neighboring DNA as a primer and polymerizes DNA from it, displacIng the RNAprimer. ADNA ligase removes nicks in the DNA by connecting the fragments together. Topoisomerase IV is required to separate the two daughter chromosomes.
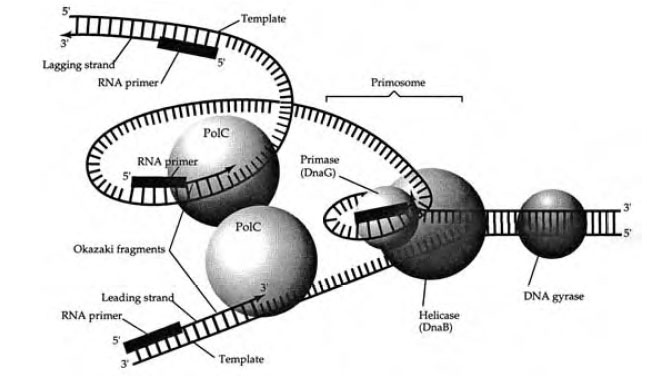 |
| Figure 3-2 Prokaryotic DNA replication. |
DNA replication in eukaryotic chromosomes generally is initiated from many origin of replication sites. Replication forks proceed in both directions from these sites. The sites that comprise yeast origins of replication are called autonomously replicating sequences (ARSs) and consist of two regions that bind a distinct set of proteins that destabilize the double helix. In one region, conserved, repeating 11-mers bind a multiprotein complex called the origin recognition complex (ORC). When proteins also bind the other region, the DNA bends by interaction of the proteins in the two regions. This distortion of the DNA promotes the separation of paired DNA strands at the origin and initiation of RNA primer synthesis.
Enzymes similar to those involved in bacterial DNA replication are found in eukaryotes. Numerous topoisomerases, helicases, and RNA polymerases have been found in eukaryotes. DNA topoisomerase II is involved in relieving positive supercoils in the DNA , whereas a helicase activity separates the two strands (Figure 3-3).
At least five different DNA polymerases have been found in eukaryotic cells. The primase (DNA polα) synthesizes lagging strand DNA . DNA polδ catalyzes leading strand synthesis. DNA polε and DNA polβ are responsible for replacIng the nucleotide gaps created when RNA primers are removed by endonucleases. A DNA ligase repairs singlestranded nicks (unconnected adjacent nucleotides) left in the DNA . DNA polγ performs DNA replication in the mitochondria.
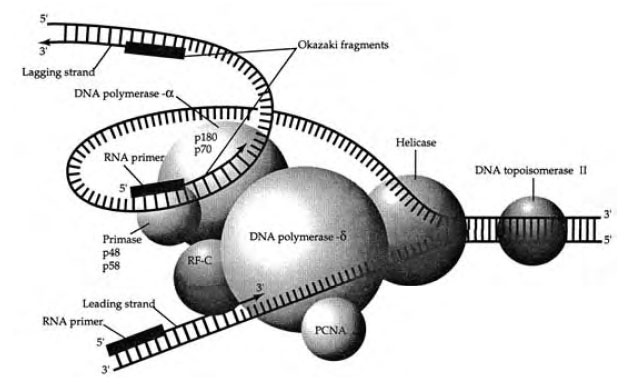 |
| Figure 3-3 Eukaryotic DNA replication. |
To complete replication of a linear chromosome, RNA primers at each end of the chromosome have to be removed and replaced by DNA . Although RNA primers can be removed by exonucleases, none of the usual DNA polymerases are able to replace the RNA without a DNA primer. An unusual type of DNA polymerase known as telomerase consists of protein and an RNAtemplate that the protein portion copies repetitively into DNA in order to extend one strand of the telomere. Thus, telomerase is responsible for maintaining the length of the chromosomes.
Notes
All the proteins and enzymes involved in replication.
Support our developers




