Symptoms of Excess
Excess K+ in plants is rare as K+ uptake is regulated strictly (64). The oversupply of K+ is not characterized by specific symptoms, but it may depress plant growth and yield (65). Excess K+ supply has an impact on the uptake of other cationic species and may thus affect crop yield and crop quality. With an increase of K+ availability in the soil, the uptake of Mg2+ and Ca2+ by oats (Avena sativa L.) was reduced (66). This action may have a negative impact for forage, where higher Mg2+ concentrations may be desirable. The relationship between K+ availability and the Mg2+ concentrations in the aerial plant parts of oats at ear emergence is shown in Figure 4.6 (66). From the graph, it is clear that the plants took up high amounts of Mg2+ only if the K+ supply was not sufficient for optimum growth. High K+ uptake may also hamper the uptake of Ca2+ and thus contribute to the appearance of bitter pit in apple (Malus pumila Mill.) fruits (67) and of blossom-end rot in tomato fruits, with strong adverse effects on fruit quality (55).The phenomenon that one ion species can hamper the uptake of another has been known for decades and is called ion antagonism or cation competition. In this competition, K+ is a very strong competitor. If it is present in a relatively high concentration, it particularly affects the uptake of Na+, Mg2+, and Ca2+. If K+ is not present in the nutrient solution, the other cationic species are taken up at high rates. This effect is shown in Table 4.4 for young barley (Hordeum vulgare L.) plants grown in solution culture (68). In one treatment with the barley, the K+ supply was interrupted for 8 days, having a tremendous impact on the Na+, Mg2+, and Ca2+ concentrations in roots and shoots as compared with the control plants with a constant supply of K+. The sum of cationic equivalents in roots and shoots remained virtually the same. This finding is explained by the highly efficient uptake systems for K+ as compared with uptake of the other cationic species. Uptake of K+ leads to a partial depolarization of the plasmalemma (the cytosol becomes less negative due to the influx of K+).
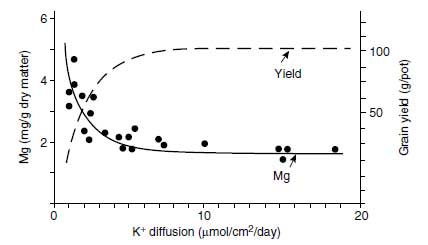 |
| FIGURE 4.6 Effect of K+ availability expressed as K+ diffusion rate in soils on the Mg concentration in the aerial plant parts of oats at ear emergence and on grain yield (Adapted from H. Grimme et al., Büntehof Abs. 4:7–8, 1974/75.) |
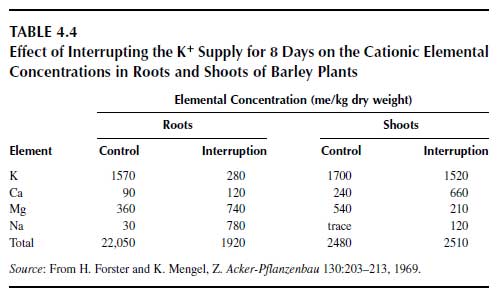 |
This depolarization reduces the driving force for the uptake of the other cationic species, which are otherwise taken up by facilitated diffusion. In the roots, the absence of K+ in the nutrient solution promoted especially the accumulation of Na+, and the shoots showed remarkably elevated Ca2+ and Mg2+ concentrations. Owing to the increased concentrations of cations except K+, the plants were able to maintain the cation-anion balance but not the growth rate. The interruption of K+ supply for only 8 days during the 2-to-3-leaf stage of barley significantly depressed growth and yield; the grain yield in the control treatment was 108 g/pot, and in the K+-interrupted treatment was 86 g/pot. This result shows the essentiality of K+ and demonstrates that its function cannot be replaced by other cationic species.
In this context, the question to what degree Na+ may substitute for K+ is of interest. The osmotic function of K+ is unspecific and can be partially replaced by Na+, as was shown for ryegrass (Lolium spp.) (69) and for rice (70). The Na+ effect is particularly evident when supply with K+ is not optimum (71). A major effect of Na+ can be expected only if plants take up Na+ at high rates. In this respect, plant species differ considerably (72). Beet species (Beta vulgaris L.) and spinach (Spinacia oleracea L.) have a high Na+ uptake potential, and in these species Na+ may substitute for K+ to a major extent. Cotton (Gossypium hirsutum L.), lupins (Lupinus spp. L.), cabbage (Brassica oleracea capitata L.), oats, potato (Solanum tuberosum L.), rubber (Hevea brasiliensisWilld. ex A. Juss.), and turnips (Brassica rapa L.) have a medium Na+ uptake potential; barley, flax (Linum usitatissimum L.), millet (Pennisetum glaucum R. Br.), rape (Brassica napus L.), and wheat have a low Na+ potential and buckwheat (Fagopyrum esculentum Moench), corn, rye (Secale cereale L.), and soybean (Glycine max Merr.) a very low Na+ uptake potential. However, there are also remarkable differences in the Na+ uptake potential between cultivars of the same species, as was shown for perennial ryegrass (Lolium perenne L.) (73).
The Na+ concentration in the grass decreased with K+ supply and was remarkably elevated by the application of a sodium fertilizer. In sugar beet, Na+ can partially substitute for K+ in leaf growth but not in root growth (74). This effect is of interest since root growth requires phloem transport and thus phloem loading, which is promoted by K+ specifically (see above). The same applies for the import of sucrose into the storage vacuoles of sugar beet (50). Also, Na+ is an essential nutrient for some C4 species, where it is thought to maintain the integrity of chloroplasts (75). The Na+ concentrations required are low and in the range of micronutrients.
Support our developers




