Metabolic Engineering of High and Low Polyunsaturated Vegetable Oils
Alteration of the a-linolenic content of seed oils is an important biotechnological target. This fatty acid is a very minor component of the seed oil of a number of crops, including corn, sunflower, peanut, and canola. α-Linolenic acid, however, accounts for nearly 10% of soybean oil and over 50% of linseed (or flax) oil. The three double bonds of this fatty acid make it particularly prone to oxidation. This is an undesirable property for food processing as the oxidation products of α-linolenic result in rancidity and reduced shelf life. Conversely, the oxidative instability of α-linolenic acid is an essential property for the use of vegetable oils such as linseed oil in drying oil applications. The free radicals generated from oxidation of α-linolenic acid-rich oils result in the enhanced polymerization (or ‘‘drying’’) of paint, ink, and other coating materials.The α-linolenic acid content of seed oils can be increased or decreased by altering the expression of genes for FAD3, the ER Δ15-linoleic acid desaturase (Table 7.4 and Fig. 7.4). As described above, FAD3 catalyzes the conversion of linoleic acid to a-linolenic acid. Transgenic expression of the A. thaliana FAD3 gene to high levels using a strong seed-specific promoter has been shown to increase the α-linolenic acid content to >50% of A. thaliana seed oil, which is comparable to the proportion found in linseed oil (Yadav et al., 1993). Downregulation of FAD3 expression in seeds can be achieved through transgenic approaches or by the generation of mutants. The development of FAD3 mutants with good agronomic performance has been particularly effective in soybean. Mutants with as little as 1–3% α-linolenic acid in their seed oil have been reported (Ross et al., 2000). These mutants do not display any significant reductions in seed yield (Ross et al., 2000). It is also notable that transgenic suppression of FAD2 genes in soybean likewise yields oils with 2–3% of a-linolenic acid. This phenotype is due to the large decrease in the linoleate substrate pool for the Δ15-linoleic acid desaturase (Buhr et al., 2002; Kinney, 1996).
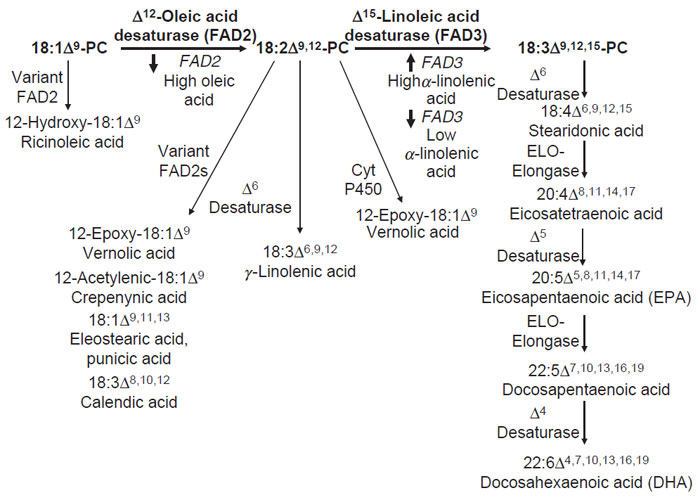 |
| FIGURE 7.4 Examples of commercially important fatty acid modification reactions that can occur in the ER of seeds. The Δ12-oleic acid desaturase or FAD2 and the Δ15-linoleic acid desaturase or FAD3 commonly occur in seeds. By up- or downregulating the expression of FAD2 and FAD3 genes, the relative levels of vegetable oil unsaturation can be altered. Variant forms of enzymes such as FAD2, cytochrome P450 monoxygenase, and cytochrome b5-fusion desaturases can be transgenically expressed in existing oilseeds to produce unusual fatty acids such as ricinoleic, vernolic, and GLAs. In addition, desaturases and ELO elongases from sources including mosses, fungi, and algae can be engineered into oilseed crops to produce the nutritionally important longchain polyunsaturated fatty acids eicosapentaenoic (EPA) and docosahexaenoic (DHA) acids. |
Support our developers




