PHA As a Bacterial Polyester
PHAs have been detected in over 90 genera of bacteria, including Gram-positive and Gram-negative species, as well as some cyanobacteria (Kim and Lenz, 2002; Steinbu¨ chel and Hein, 2001). While the majority of PHAs are composed of monomers of R-(—)-3-hydroxyalkanoic acid ranging from 3 to 16 carbons in length (C3–C16) (Fig. 8.3), some PHAs can also incorporate 4-, 5-, or 6-hydroxy acids (Steinbu¨ chel and Valentin, 1995). Nearly 150 different hydroxyacids have been found incorporated in bacterial PHAs, with the major diversity being found in the length and the presence of functional groups in the side–chains of the polymer (Steinbu¨ chel and Valentin, 1995). Although some of these monomers have been found in PHA produced by bacteria in their natural environment, a larger fraction of monomers have been incorporated into PHA following growth of bacteria in artificial media containing exotic sources of carbon, such as fatty acids with double or triple bonds.Bacteria synthesizing PHAs have been broadly subdivided in two groups. One group, including the bacterium Ralstonia eutropha, produces short-chain-lengthPHA (SCL-PHA) containing monomers ranging from 3 to 5 carbons in length, while a second group, including a number of Pseudomonads, synthesizes medium chain length-PHA (MCL-PHA) containing monomers ranging from 6 to 16 carbons in length. This division
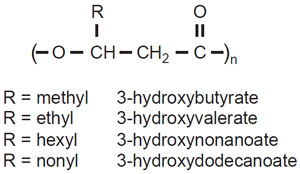 |
| FIGURE 8.3 Chemical structure of polyhydroxyalkanoate. The monomers can range from 3 to 16 carbons in length, depending on the size of the pendant R group. |
between SCL- and MCL-PHA is mainly determined by the substrate specificity of the PHA synthase responsible for the polymerization of the substrate R-3-hydroxyacyl-CoA to form PHA. This division between SCL- and MCL-PHA is however not strict, since several bacteria have been found that can synthesize a ‘‘hybrid’’ PHA that can include monomers from 4 to 8 carbons (Fukui and Doi, 1997).
A number of enzymes and metabolic pathways have been implicated in the synthesis of a spectrum of PHAs in bacteria. In this chapter, we wish to focus only on the pathways that have been successfully transferred in plants. The readers are referred to several excellent reviews to learn more on various aspects of bacterial PHA, including biochemical synthesis and application of PHAs (Anderson and Dawes, 1990; Braunegg et al., 1998; Steinbü chel, 1991; Steinbü chel and Fü chtenbusch, 1998; Steinbü chel and Lu¨ tke-Eversloh, 2003; Steinbü chel and Schlegel, 1991; Sudesh and Doi, 2000; van der Walle et al., 2001).
Support our developers




