Cryptophyta
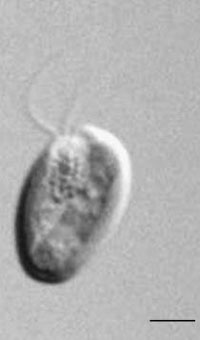
FIGURE 1.36 Unicell of Cryptomonas sp. (Bar: 6 µm.)
The unicellular flagellates belonging to the division Cryptophyta are asymmetric cells dorsiventrally
constructed (Figure 1.36). They bear two unequal, hairy flagella, subapically inserted, emerging
from above a deep gullet located on the ventral side of the cell. The wall of this gullet is lined
by numerous ejectosomes similar to trichocysts. Cryptophytes are typically free-swimming in
freshwater and marine habitats; palmelloid phases can also be formed, and some members are
known to be zooxanthellae in host invertebrates or within certain marine ciliates. Cryptophyta
possess only chlorophylls a and c
2. Phycobilins are present in the thylakoid lumen rather than in
phycobilisomes. The chloroplasts, one or two per cell, are surrounded by a fold of the endoplasmic
reticulum.
In the space between these membranes a peculiar organelle, the nucleomorph, is located.
This organelle can be interpreted as the vestigial nucleus of the red algal endosymbiont that gave
rise to the chloroplasts of the Cryptophyta. Thylakoids are arranged in pairs, with no girdle lamellae.
The pyrenoid projects out from the inner side of the chloroplast. The chloroplast DNA is condensed
in small nucleoids scattered inside the chloroplast. The reserve polysaccharide accumulates in the periplastidial space as starch granules. Sometimes an eyespot formed by spherical globules is
present inside the plastid, but it is not associated with the flagella. The cell is enclosed in a stiff,
proteinaceous periplast, made of polygonal plates. Most forms are photosynthetic, but heterotrophic
nutrition also occurs. The primary method of reproduction is simply by longitudinal cell division,
but sexual reproduction has recently been documented.
FIGURE 1.36 Unicell of Cryptomonas sp. (Bar: 6 µm.)
DINOPHYTA
The members of this division are typical unicellular flagellates (Figure 1.37) but can be also nonflagellate,
ameboid, coccoid, palmelloid, or filamentous. Dinoflagellates have two flagella with
independent beating pattern, one training and the other girdling that confers characteristic rotatory
swimming whirling motion. Flagella are apically inserted (desmokont type) or emerge from a
region close to the midpoint of the ventral side of the cell (dinokont type). Most dinoflagellates
are characterized by cell-covering components that lie beneath the cell membrane. Around the
cell there is a superficial layer of flat, polygonal vesicles, which can be empty or filled with cellulose plates. In dinokont type dinoflagellates, these thecal plates generally form a bi-partite armor, consisting
of an upper, anterior half and a lower, posterior half, separated by a groove known as cingulum
where the transversal flagellum is located (Figure 1.38). A smaller groove, the sulcus,
extends posteriorly from the cingulum, and hosts the longitudinal flagellum. The two flagella
emerge from a pore located at the intersection of the two grooves. Very often they are important
components of the microplankton of freshwater and marine habitats.
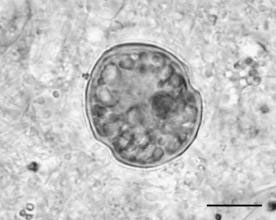
FIGURE 1.37 A marine dinoflagellate. (Bar: 30 µm.)
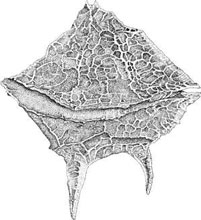
FIGURE 1.38 Dorsal view of Gonyaulax sp., a brackish water dinoflagellate.
Though most are too large
(2–2000 mm) to be consumed by filter feeders, they are readily eaten by larger protozoa, rotifer,
and planktivorous fishes. Some Dinoflagellates are invertebrate parasites, others are endosymbionts
(zooxanthellae) of tropical corals. Dinoflagellates possess chlorophylls a, b, c1, and c2, fucoxanthin,
other carotenoids, and xanthophylls such as peridinin, gyroxanthin diester, dinoxanthin, diadinoxantin,
and fucoxanthin. The chloroplasts, if present, are surrounded by three membranes. Within the
chloroplasts the thylakoids are for the most part united in a stack of three. The chloroplast DNA is
localized in small nodules scattered in the whole chloroplast, with typical pyrenoids. A really
complex photoreceptive system is present in the dinophytes such as Warnowia polyphemus,
Warnowia pulchra, or Erythropsidinium agile consisting of a “compound eye” composed of a
lens and a retinoid. Most dinoflagellates are distinguished by a dinokaryon, a special eukaryotic
nucleus involving fibrillar chromosomes that remain condensed during the mitotic cycles. The principal
reserve polysaccharide is starch, located as grains in the cytoplasm, but oil droplets are present
in some genera.
At the surface of the cell there are trichocysts which discharge explosively when
stimulated. Besides photoautotrophy, dinoflagellates exhibit an amazing diversity of nutritional
types because about half of the known species lack plastids and are therefore obligate heterotrophic.
Some are notorious for nuisance blooms and toxin production, and many exhibit bioluminescence.
Dinophyceae have generally a haplontic life history.




