Cyanophyta and Prochlorophyta
All blue-green algae (Figure 1.25) and prochlorophytes (Figure 1.26) are non-motile Gramnegative eubacteria. In structural diversity, blue-green algae range from unicells through branched and unbranched filaments to unspecialized colonial aggregations and are possibly the most widely distributed of any group of algae. They are planktonic, occasionally forming blooms in eutrophic lakes, and are an important component of the picoplankton in both marine and freshwater systems; benthic, as dense mats on soil or in mud flats and hot springs, as the “black zone” high on the seashore, and as relatively inconspicuous components in most soils; and symbiotic in diatoms, ferns, lichens, cycads,sponges, and other systems. Numerically these organisms dominate the ocean ecosystems. There are approximately 1024 cyanobacterial cells in the oceans. To put that in perspective, the number of cyanobacterial cells in the oceans is two orders of magnitude more than all the stars in the sky.
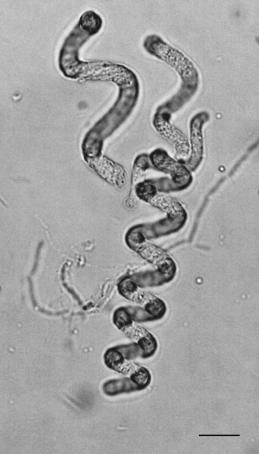
FIGURE 1.25 Trichome of Arthrospira sp. (Bar: 20 µm.) (Bar: 0.5 µm.)
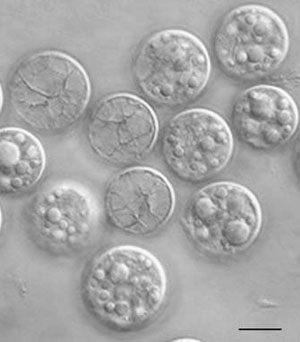
FIGURE 1.26 Cells of Prochloron sp. (Bar: 10 µm.)
Pigmentation of cyanobacteria includes chlorophyll a, blue and red phycobilins (phycoerythrin, phycocyanin, allophycocyanin, and phycoerythrocyanin), and carotenoids. These accessory pigments lie in the phycobilisomes, located in rows on the outer surface of the thylakoids. Their thylakoids, which lie free in the cytoplasm, are not arranged in stacks, but singled and equidistant, in contrast to prochlorophytes and most other algae, but similar to Rhodopyta and Glaucophyta.
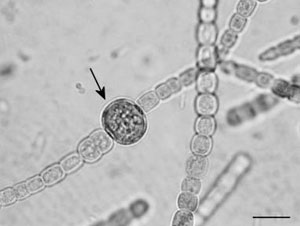
FIGURE 1.27 Heterocyst (arrow) of Anabaena azollae. (Bar: 10 µm.)
The reserve polysaccharide is cyanophycean starch, stored in tiny granules lying between the thylakoids. In addition, these cells often contain cyanophycin granules, that is, polymer of arginine and asparagine. Some marine species also contain gas vesicles used for buoyancy regulation. In some filamentous cyanobacteria, heterocysts and akinetes are formed. Heterocysts are vegetative cells that have been drastically altered (loss of photosystem II, development of a thick, glycolipid cell wall) to provide the necessary anoxygenic environment for the process of nitrogen fixation (Figure 1.27). Some cyanobacteria produce potent hepato- and neurotoxins.
FIGURE 1.27 Heterocyst (arrow) of Anabaena azollae. (Bar: 10 µm.)
Prochlorophytes can be unicellular or filamentous, and depending on the filamentous species, they can be either branched or unbranched. They exist as free-living components of pelagic nanoplankton and obligate symbionts within marine didemnid ascidians and holothurians, and are mainly limited to living in tropical and subtropical marine environments, with optimal growth temperature at about 24°C.
Prochlorophytes possess chlorophylls
a and
b similar to euglenoids and land plants, but lack phycobilins, and this is the most significance difference between these and cyanobacteria; other pigments are β-carotene and several xanthophylls (zeaxanthin is the principal one). Their thylakoids, which lie free in the cytoplasm, are arranged in stacks. Prochlorophytes have a starch-like reserve polysaccharide. These prokaryotes contribute a large percentage of the total organic carbon in the global tropical oceans, making up to 25–60% of the total chlorophyll a biomass in the tropical and subtropical oceans. They are also able to fix nitrogen, though not in heterocysts.
Both blue-green algae and prochlorophytes contain polyhedral bodies (carboxysomes) containing RuBisCo (ribulose bisphospate carboxylase/oxygenase, the enzyme that converts inorganic carbon to reduced organic carbon in all oxygen evolving photosynthetic organisms), and have similar cell walls characterized by a peptoglycan layer. Blue-green algae and Prochlorophytes can be classified as obligate photoautotrophic organisms. Reproduction in both divisions is strictly asexual, by simple cell division of fragmentation of the colony or filaments.




