Type 3: Cell Surface with Additional Intracellular Material in Vesicles
According to Dodge and Crawford (1970), the amphiesma construction falls into eight reasonably distinct categories:
- (1) simple membrane underlain by a single layer of vesicles 600–800 nm in length, rather flattened, circular, or irregular in shape, with a gap of at least 40 nm between adjacent vesicles that may contain dense granular material; beneath the vesicles are parallel rows of microtubules which lie in groups of three; this simple arrangement is present in Oxyrrhis marina;
- (2) simple membrane underlain by closely packed polygonal (generally hexagonal) vesicles 0.8–1.2 µm in length, frequently containing fuzzy material; these vesicles and the cell membrane are occasionally perforated by trichocyst pores; beneath the vesicles lie microtubules in rows of variable number; this type of amphiesma has been found in Amphidinium carteri;
- (3) as in category (2), but with plug-like structures associated with the inner side of the vesicles; these plugs are cylindrical structures 120 nm long, and are arranged in single lines between single or paired microtubules; an example of this arrangement is present in Gymnodinium veneficum;
- (4) as in category (2), but with thin (about 20 nm) plate-like structure in the flattened vesicles; this amphiesma characterizes Aureodinium pigmentosum;
- (5) in this group the vesicles contain plates of medium thickness (60 nm), which slightly overlap; in Woloszynskia coronata the plates are perforated by trichocyst pores;
- (6) the plates are thicker (up to 150 nm), reduced in number with a marked diversity of form; each plate has two or more sides bearing ridges and the remaining sides have tapered flanges; where the plates join, one plate bears a ridge and the opposite bears a flange; Glenodinium foliaceum belongs to this category;
- (7) the plates can be up to 25 µm large and up to 1.8 mm thick; they bear a corrugated flange on two or more sides, and a thick rim with small projections on the opposing edges; these plates may overlap to a considerable extent, and their surface may be covered by a pattern of reticulations; a distinctive member of this category is Ceratium sp.;
- (8) amphiesma consisting of two large plates, with one or more small plates in the vicinity of the flagellar pores at the anterior end of the cell; plates can be very thin and perforated by two or three simple trichocyst pores as in Prorocentrum nanum, or thick and with a very large number (up to 60) of trichocyst pores as in Prorocentrum micans (Figure 2.18).
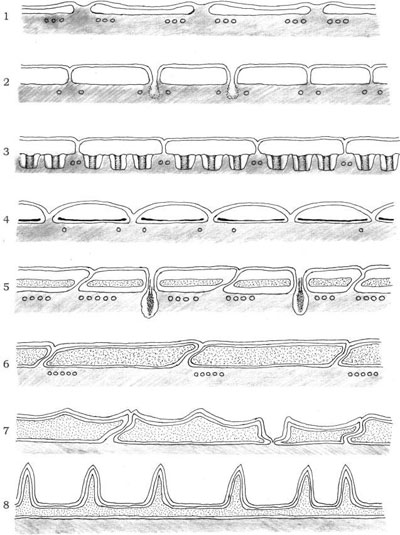
FIGURE 2.18 Diagram of the eight distinct categories of the dinoflagellate amphiesma.
The arrangement of thecal plates is termed tabulation, and it is of critical importance in the taxonomy of dinoflagellates. Tabulation can also be conceived of as the arrangement of amphiesmal vesicles with or without thecal plates. The American planktologist and parasitologist Charles Kofoid developed a tabulation system allowing reference to the shape, size, and location of a particular plate; plates were recognized as being in series relative to particular landmarks such as the apex, cingulum (girdle), sulcus. His formulas (i.e., the listing of the total number of plates in each series) were especially useful for most gonyaulacoid and peridinioid dinoflagellates.
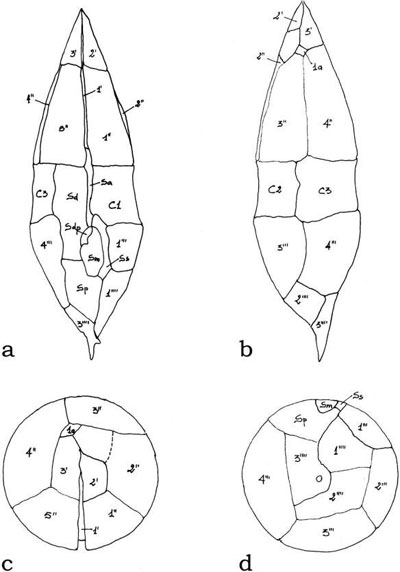
FIGURE 2.19 Line drawings of the thecal plate patterns of Lessardia elongata with the corresponding numeration. Ventral view (a), dorsal view (b), apical view (c), and antiapical view (d).
Support our developers




