Type I
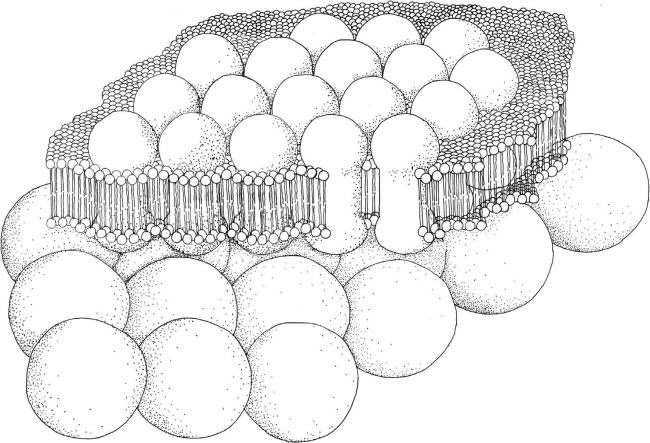
FIGURE 2.65 Schematic drawing of Type I photoreceptor system.
A single layer of photoreceptor molecules is present inside the whole cell membrane, or located in just the patch of membrane that covers the eyespot when present (Figure 2.65). Lacking the eyespot, the whole algal body performs the shading function. This means that this type of photoreceptive system could be not readily visible.
FIGURE 2.65 Schematic drawing of Type I photoreceptor system.
In Cyanophyta, phototactic orientation has been described in Anabaena variabilis, Pseudoanabaena sp., and Phormidium, although no defined structure for light sensing have been so far detected. The first identification of a complex photoreceptive system leading to the evidence of a photoreceptive protein was performed in Leptolyngbya sp. This deep red cyanobacterium lives in Roman hypogea at extremely low light intensity (10
13 photons m
-2 sec
-1). It possesses an orange eyespot at the tip of the apical cell of the trichome. Electron microscopy revealed that this eyespot is characterized by osmiophilic globules of about 100 nm in diameter arranged in a peripheral cap extending 2–3 µm from the apex and with a possible layered pattern. Microspectrophotometric analysis of the tip of the apical cell of
Leptolyngbya trichomes revealed a complex absorption spectrum with two main bands. The band centered at 456 nm is due to the absorption of the carotenoid present in the eyespot, whereas the band centered at 504 nm can be assigned to rhodopsin-like molecules packed in the plasma membrane of the tip of the apical cell.
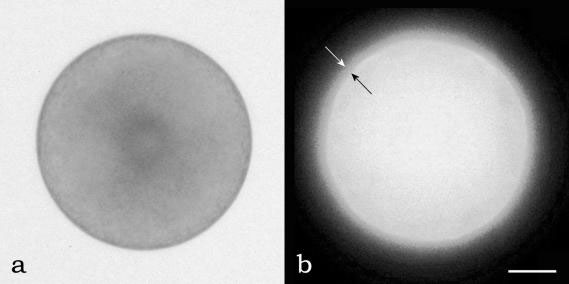
FIGURE 2.66 A S. compressa zygote in bright field microscopy (a), and under fluorescence microscopy (b) labeled with anti-rhodopsin antibody. The two arrows point to the cell membrane layer of the cell in which photoreceptive protein are located. (Bar: 2 µm.)
In Heterokontophyta data exist indicating that the photoreceptor molecules are present inside the cell membrane of zygotes of the fucoid brown algae,
Fucus sp. and
Silvetia compressa. Experimental work confirmed this localization in
S. compressa (Figure 2.66a), where a rhodopsin-like protein was identified in the zygote membrane (Figure 2.66b). Because no eyespot can be detected in these algal stages, the shading function is assigned to the whole cell body.
FIGURE 2.66 A S. compressa zygote in bright field microscopy (a), and under fluorescence microscopy (b) labeled with anti-rhodopsin antibody. The two arrows point to the cell membrane layer of the cell in which photoreceptive protein are located. (Bar: 2 µm.)
In the Haptophyta, the eyespot is present only in some species of the order Pavlovales. It consists of a single layer of globules situated at the anterior end of one of the chloroplast, beneath the posteriorly directed flagellum, at the level of its emersion from the cell. In these algae the photoreceptor has not been yet localized, but we can presume it is positioned inside the membrane in close association with the eyespot.
In the Cryptophyta, the presence of an eyespot is limited to a small number of species belonging to the genus
Chroomonas. The eyespot is situated in the center of the cell, within a conical lobe of the chloroplast. It consists of a single layer of about 35 closely packed globules, attached to the chloroplast envelope and the endoplasmic reticulum. Also in these algae the photoreceptive proteins should be located inside the plasma membrane overlaying the eyespot.
In the Dinophyta, the eyespot is chloroplastic in
Peridiniun sp., consisting of a layer of globules under the chloroplast envelope, situated behind the longitudinal sulcus, and truly extraplastidic in
Woloszynskia coronata, where it consists of an irregular cluster of globules located beneath the sulcus, and immediately adjacent to the subthecal microtubules. In the latter, neither connection with the chloroplast nor membranes surrounding the eyespot are present.
Glenodinium foliaceum and
Peridinium balticum possess another type of eyespot. It is a roughly triangular body situated behind the sulcus and it is an independent structure bounded by a three-membrane envelope. Basically, there are two layers of pigmented globules, separated by a vesicle of granular material. The eyespot can fold back upon itself, making more layers. In
Amphidinium lacustre the eyespot is an elongated structure located along the right edge of the sulcus. Its color is a shade of greenish-yellow rather than the red-orange, commonly found in eyespot; it consists of up to six flat rows of brick-like units, each row contained in a vesicle bounded by a unit membrane. No data exist on the photoreceptor location in these dinoflagellates, but the assumption is the same made for the Haptophyta and Cryptophyta, that is, photoreceptive proteins must be located inside the plasma membrane close to the eyespot. A separate case is that of
Alexandrium hiranoi and
Gymnodinium mikimotoi. Both dinoflagellates show phototactic responses but lack a detectable eyespot, hence the shading function is performed by the cell body.
In Chlorophyta such as
Haematococcus sp.,
Spermatozopsis sp., and
Dunaliella sp., the eyespot is situated on one side of the cell, sometimes slightly protruding beyond the cell surface, as in the gametes of
Ulva; its area can range from about 0.3 to 10 µm
2. The globules range from 80 to 190 nm in diameter and their number varies from 30 to approximately 2000. The most common organization consists of a single layer of closely packed globules lying between the outermost thylakoid and the two-layered chloroplast membrane. Additional layers of globules can be present underneath the first layer, individual layers subtended or not by a single thylakoid. In most species the globules show a hexagonal packing, which enable the highest possible packing density. In both
Pandorina and
Volvox colonies, the eyespot of cells in the anterior of the
colony are larger than those of the posterior, consisting of up to nine layers, marking the occurrence of some degree of colony polarity. The photoreceptor of Chlamydomonas can be considered the model of Type I photoreceptor. It consists of an extensive two-dimensional patch of photosensitive proteins, identified as rhodopsin-like protein, localized in the plasma membrane overlying the
eyespot. The layered structure of the shading organelle in this type of photoreceptor works as a quarter wave interference reflector that reflects the impinging light toward the photoreceptor, in order to increase detectability of the light signal.




