Isolation of DNA to be Cloned
Keeping in view the aim of the experiment, DNA molecule is either enzymatically fragmented or may be procured from cDNA or gene banks. Getting a cDNA fragment of its known function or characterizing its sequence is very difficult. A cDNA fragment is prepared directly by using mRNA as template. It follows many biochemical methods. Necessary enzymes with their restriction sites and cleaved products required during the isolation of DNA have been described earlier, and the important enzymes are given in Table 3.2. It is much difficult to isolate a specific mRNA if it is in low amount in the cell. The majority of mRNA sequence in eukaryotic cells contain a long polyadenylated tract (
i.e. about 100A residues) at their 3' termini (Kates, 1970).
By this virtue they can be separated from other types of RNAs. Therefore, mRNA binds with an oligo-dT cellulose affinity column or poly-U sepharose from which it can be eluted. Commercial preparations of both these substrates are now available. The mRNA preparations can be enriched for the desired molecules by fractionation according to their size by using sucrose density gradient centrifugation (Boffey, 1987). Preparation of cDNA is shown in Fig. 2.13. Reverse transcriptase is required for the synthesis of DNA copy of an mRNA molecule in the presence of oligo-dT primer. Cellular DNA and total RNA inhibit reverse transcriptase activity. Therefore, it is necessary that mRNA must be in pure form before cloning.
The oligo-dT segments contain between 10-20 nucleotides in length that hybridize to the poly-A tract on mRNA. The oligo-dT primer ensures the initiation of cDNA synthesis at 3 terminus of RNA in poly-A tract. The conditions that optimize the synthesis of cDNA are:
(i) The use of high level of deoxynucleoside triphosphates
i.e. dATP, dCTP, dGTP, dTTP (Kacian and Myers, 1976).
(ii) Addition of sodium pyrophosphate or ribonucleoside triphosphates (Kacian and Myers, 1976),
(iii) Omission of monovalent cations coupled with evaluation of the reaction temperature to 46°C (Monahan
et al, 1976).
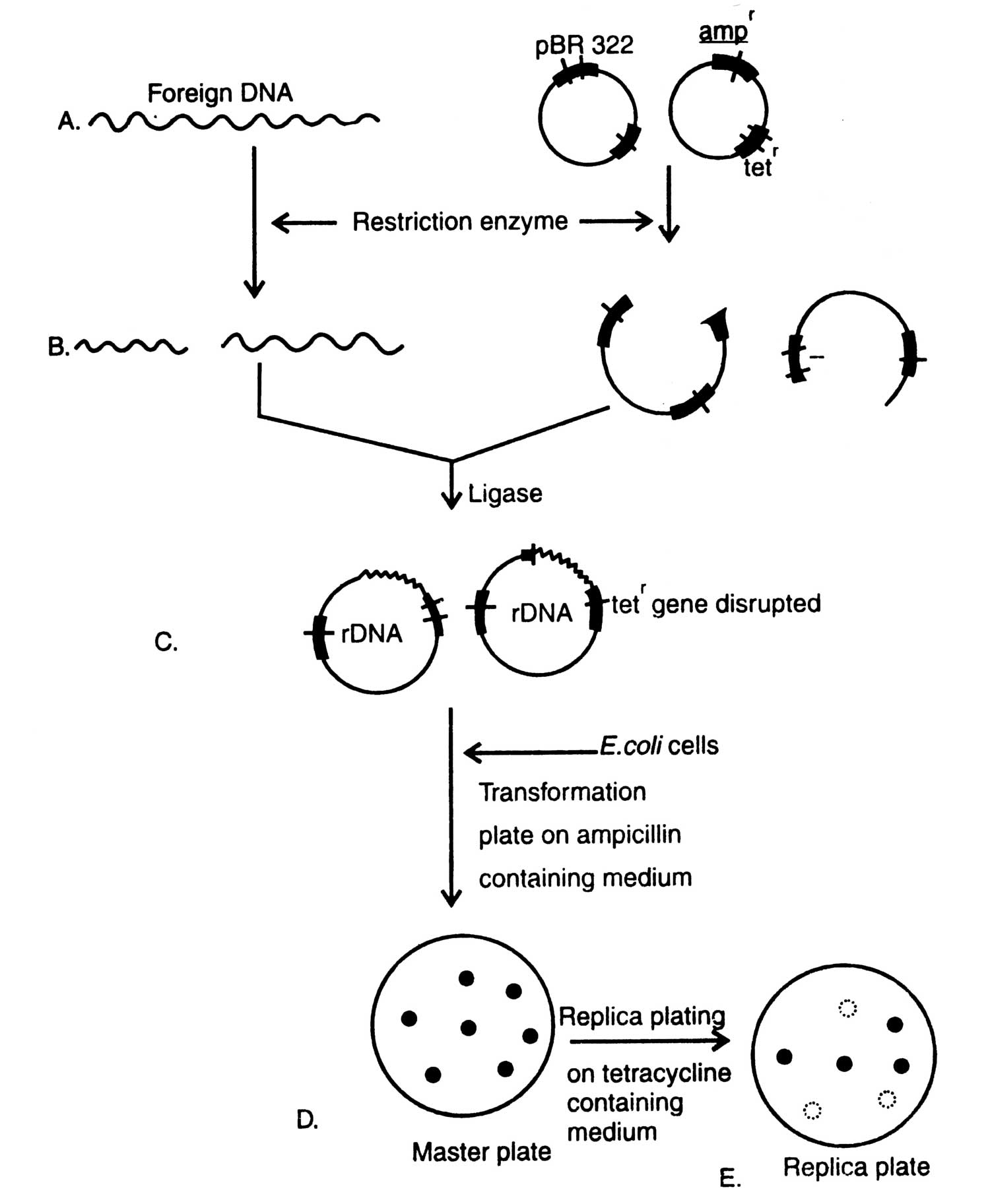
Fig. 4.1. Diagrammatic presentation of gene cloning.
This results in the synthesis of mRNA-cDNA hybrid. On addition of terminal transferase and dCTP the oligo-dC tails at 3' termini of both strands are formed and the residues of cDNA become curved. The curved tail forming a loop like structure is known as hairpin loop (Leis and Hurritz., 1972). Upon hydrolysis, by using alkali, the mRNA-cDNA strands are separated into single strand. Now cDNA acts as a template for the synthesis of double stranded cDNA in the presence of DNA polymerase I or AMV (Avian myeloblastosis virus) reverse transcriptase.
The hairpin acts as a primer for elongation. This results in the synthesis of a double stranded copy of the cDNA which contains one closed end (at 5' end of mRNA) and the other open end (at 3'end of mRNA) SI nuclease is used to cleave hairpin loops and results in double stranded cDNAs which are further used in gene cloning experiments.
The double stranded cDNA is identical to the gene which codes for mRNA( only in prokaryotic genes). However, the majority of eukaryotic genes contain introns (the non coding regions) which interrupt their encoding sequences. Methods of removing cDNA in eukaryotes have been described earlier
(see 3.7). cDNA can be radioactively labeled by nick translation and used as a hybridization probe for the identification of DNA fragments containing the gene. By using these procedures a cDNA clone bank is built up
(see 3.6). Procedure of gene cloning is shown in Fig. 4.1.




