Gene vs allele : A new concept of allelomorphism
Pseudoalleles and complex loci
 Earlier in this section, we discussed criteria for allelism, which included lack of recombination and lack of complementation between two mutant alleles of the same gene. In earlier sections, we also defined allelomorphs (or alleles) as alternative forms of same gene. This classical concept of allelomorphism needed change in view of intensive work done after 1940, initially in Drosophila and later in several other organisms.
Earlier in this section, we discussed criteria for allelism, which included lack of recombination and lack of complementation between two mutant alleles of the same gene. In earlier sections, we also defined allelomorphs (or alleles) as alternative forms of same gene. This classical concept of allelomorphism needed change in view of intensive work done after 1940, initially in Drosophila and later in several other organisms.
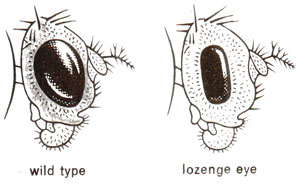
In Drosophila, a recessive sex linked mutant called lozenge (lz)is responsible for smaller, darker and more elliptically shaped eyes (Fig. 14.10). A number of mutants at this locus were discovered and could be identified due to severity of effect on eye and also due to other phenotypic effects. Ordinarily a fly heterozygous for two lozenge mutant alleles (lz1/lz2)will not yield wild type in its progeny, because wild allele is absent. However in 1940, Oliver discovered wild type flies in progeny of such heterozygotes, with a frequency of one in several thousands. This frequency was still much higher than expected on the basis of spontaneous mutation rates known. In view of this, the results could not be easily explained and the lozenge locus was then thoroughly studied by M.M. Green, a student of Oliver.
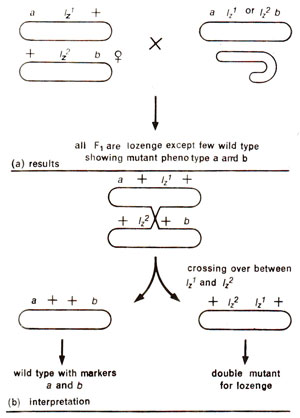
Following Sturtevant's technique with Bar locus, Green marked the lozenge locus on either side by marker genes to confirm if the wild type appeared due to recombination. If wild type was the result of recombination, Green expected that marker genes would also recombine. In his experiments, Green observed that not only wild type flies appeared for lozenge, but the marker genes a and b also recombined suggesting that lz1 and Iz2 could recombine (Fig. 14.11). This suggested that two mutants may be separated by a distance within the gene.
Later E.B. Lewis, in 1951 reported results of one of his experiments, where from a cross of apricot eyed and white eyed flies (apricot is an allele of white eye locus), he obtained F1 having intermediate eye colour. In F2, he had expected segregation only for apricot and white, but to his surprise, he recovered at a very low frequency wild type also as earlier shown by Oliver and Green in case of lozenge locus.
In both examples of lozenge and eye colour, mutant alleles apparently recombined and therefore proved to be nonallelic according to the classical concept of allelomorphism described earlier. Since these alleles behaved as non-allelic, Lewis preferred to call them pseudoalleles and the phenomenon as pseudoallelism. Pontecorvo later however argued that since our concept of allelism has changed and that the classical concept of alleles has been modified in view of this work, it is needless to call this phenomenon as pseudoallelism and the elements as pseudoalleles. He, instead, suggested that the phenomenon be called Lewis effect and the mutant elements be called alleles as earlier.
Position pseudoallelism or cis-trans effect In above experiments of Green and Lewis we tried to demonstrate that intragenic interallelic recombination can take place leading to appearance of wild type (in a very small frequency) in the progeny of a F1 heterozygote for two mutant alleles. If two mutant alleles occupy different positions, showing close linkage rather than same position, then one may expect that F, heterozygote should have a genotype lz1+/+lz2 or apr+/+w and, therefore, should express wild phenotype. Since F1 did not exhibit wild phenotype, we may like to examine the second criterion for allelism i.e. lack of complementation. In view of this criterion, we may explain the absence of wild type in F1 due to lack of complementation. Such a lack of complementation is due to position effect, which can be understood by studying two arrangements of mutant alleles.
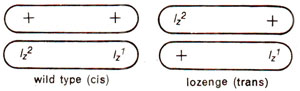
For instance in heterozygote for lz1 and lz2, two arrangements are possible (Fig. 14.12), one with both mutant alleles on same chromosome and their wild counterparts on the other homologue (cis), the other with two mutant alleles on two different homologues (trans). For lozenge locus (lz1 and lz2)cis and trans arrangements are shown in Figure 14.12. It is also indicated that in cis position they will express wild type, and in trans configuration, the mutant phenotype. This may presumably be due to position effect, where a mutant allele does not allow adjacent region to express wild phenotype. This difference in phenotypes due to cis and trans arrangements is known as cis-trans effect.
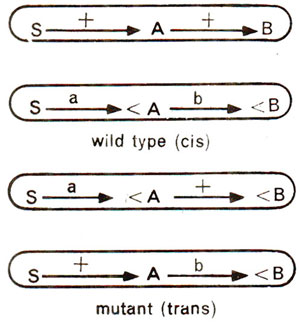
An explanation for cis-trans effect was given by E.B. Lewis, who believed in sequential nature of gene product synthesis. According to him, if both mutant alleles are on one chromosome, the other chromosome will be normal and will be able to produce the end product. Such a situation is found in cis arrangement. But in trans arrangement, sequence of steps involved in synthesis will be interrupted due to mutations on either of the two homologous chromosomes, thus leading to a mutant phenotype (Fig. 14.13).
Fig. 14.10. Wild and lozenge eye phenotypes in Drosophila.
Fig. 14.11. Results and interpretation of Green's experiment in Drosophila where female Iz1 /Iz2 was crossed with male Iz |— or lz2|—.
Fig. 14.12. Cis and trans configurations of heterozygotes involving two alleles lz1 and lz2 at lozenge eye locus in Drosophila.
Fig. 14.13. Possible explanation at the molecular level in terms of synthesis of polypeptides in cis and trans configurations.
Following Sturtevant's technique with Bar locus, Green marked the lozenge locus on either side by marker genes to confirm if the wild type appeared due to recombination. If wild type was the result of recombination, Green expected that marker genes would also recombine. In his experiments, Green observed that not only wild type flies appeared for lozenge, but the marker genes a and b also recombined suggesting that lz1 and Iz2 could recombine (Fig. 14.11). This suggested that two mutants may be separated by a distance within the gene.
Later E.B. Lewis, in 1951 reported results of one of his experiments, where from a cross of apricot eyed and white eyed flies (apricot is an allele of white eye locus), he obtained F1 having intermediate eye colour. In F2, he had expected segregation only for apricot and white, but to his surprise, he recovered at a very low frequency wild type also as earlier shown by Oliver and Green in case of lozenge locus.
Position pseudoallelism or cis-trans effect In above experiments of Green and Lewis we tried to demonstrate that intragenic interallelic recombination can take place leading to appearance of wild type (in a very small frequency) in the progeny of a F1 heterozygote for two mutant alleles. If two mutant alleles occupy different positions, showing close linkage rather than same position, then one may expect that F, heterozygote should have a genotype lz1+/+lz2 or apr+/+w and, therefore, should express wild phenotype. Since F1 did not exhibit wild phenotype, we may like to examine the second criterion for allelism i.e. lack of complementation. In view of this criterion, we may explain the absence of wild type in F1 due to lack of complementation. Such a lack of complementation is due to position effect, which can be understood by studying two arrangements of mutant alleles.
Fig. 14.10. Wild and lozenge eye phenotypes in Drosophila.
Fig. 14.11. Results and interpretation of Green's experiment in Drosophila where female Iz1 /Iz2 was crossed with male Iz |— or lz2|—.
Fig. 14.12. Cis and trans configurations of heterozygotes involving two alleles lz1 and lz2 at lozenge eye locus in Drosophila.
Fig. 14.13. Possible explanation at the molecular level in terms of synthesis of polypeptides in cis and trans configurations.
An explanation for cis-trans effect was given by E.B. Lewis, who believed in sequential nature of gene product synthesis. According to him, if both mutant alleles are on one chromosome, the other chromosome will be normal and will be able to produce the end product. Such a situation is found in cis arrangement. But in trans arrangement, sequence of steps involved in synthesis will be interrupted due to mutations on either of the two homologous chromosomes, thus leading to a mutant phenotype (Fig. 14.13).
Support our developers




