
Fig. 37.15. Heavy and light chains and their association to generate different V (variable) domains and C (constant) domains in immunoglobulin protein.
Antibody molecules, called immunoglobulins, are coded by genes, which undergo DNA rearrangements during cell differentiation, thus suggesting that although a genome usually remains constant in different cells, it does undergo a change with regard to immunoglobulin genes in mammals. Immunoglobulins are produced by cells originating in bone marrow
(B-cells), and later differentiating into lymphocytes. These may differentiate into several clones,
producing different antibodies, but each clone producing only one antibody. For this work on the rearrangement of DNA sequences that results in a large variety of sequences for production of antibodies, S.
Tonegawa of MIT (USA) was awarded the 1987 Nobel Prize for Physiology and Medicine. This work will be briefly described in this section.
Structure of immunoglobulin chains. Antibody molecules (immunoglobulins, abbreviated as I
g) are tetrameric, each containing two identical
'heavy' (H) chains of about 500 amino acid residues, and two identical
light (L) chains of about 250 amino acid residues (Fig. 37.15). In man, mouse, rabbit and other mammals there are seven kinds of heavy chains (alpha = α; gamma = γ
1, γ
2a, γ
2b, γ
3; delta = δ; mu = μ) and two kinds of light chains (kappa = κ and, lambda = λ)
. Ig molecules are named according to the type of H chain, which they contain (e.g.
IgG has gamma chains,
IgM has mu chains and
IgA has alpha chains).
Fig. 37.15. Heavy and light chains and their association to generate different V (variable) domains and C (constant) domains in immunoglobulin protein.
Each I
g chain consists of two distinct parts (i) C-terminal constant part or
C-region and (ii) N-terminal extremely variable part or
V-region. For each C-region (C
α, C
μ, C
λ, etc.), there can be several V sequences. Separate set of V sequences are known for heavy chains (V
H) and light chains (V
κ = for kappa and V
λ for lambda). The V
H, V
κ and V
λsequences have fair degree of homology with only some variable portions.
Ig gene in germline and differentiated cells. By using cDNA as probe, it could be shown that C- and V-coding sequences were closer in the antibody producing cells, but separated in the germ line. This was predicted because, same C-region was found combined with different V-regions in the same individual. Later experiments showed that C—C splicing at the DNA level is responsible for generating the coding genes for both light and heavy chains. Although, the genes for immunoglobulin molecule may be found on more than one chromosome, but cutting and rejoining of DNA takes place only within a chromosome and not across chromosomes. In addition to C and V regions, both light and heavy chains have another region called J (junction of joining sequences). In heavy chain, in addition to C, J and V regions, another short sequence D (diversity) is also found.
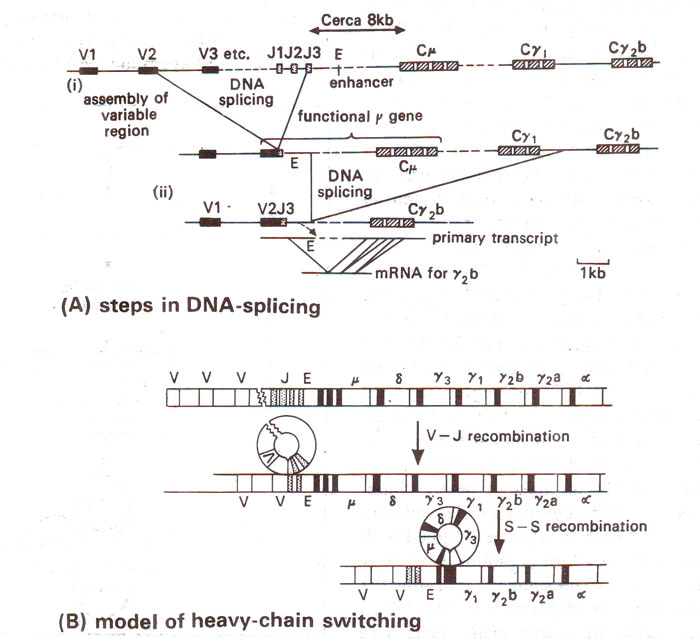
Fig. 37.16. (A) structure of immunoglobulin gene and the steps involved in DNA rearrangements in different regions. (B) a model explaining DNA rearrangements in heavy chain switching.
V-regions through DNA splicing. We will discuss DNA splicing only for heavy chain as an example. There are following two rearrangements in this case; (i) the first, generating the DNA sequence coding for V
H and (ii) the second leading to association of specific V
H with a definite C
H sequence. Using probes in different kinds of cells, steps involved in rearrangements leading to association of different V regions could be worked out as shown in Figure 37.16. In I
gM sequence, it is shown that in embryo or germ line cells, V
H and C
H regions have a small region of J sequence. Between V and J sequences, presence of D sequence was also shown. Although there is diversity in V, J and D sequences (C is constant) but variation is also provided by the manner in which DNA splicing is brought about.
V-C association through DNA splicing. Second phase of DNA splicing involves association of a V segment with C segment. It has been shown that the same V segment can attach with different C-regions, thus providing for
H-chain class switching. This is achieved by successive deletions of DNA-leading to the production of I
gM in primary response and I
gG and I
gA as secondary response. I
gM has all kinds of heavy chains; while in I
gG
3, segment C is missing, in I
gG
1, C
μ and C
4λ3 are missing. Further deletions may occur in other cases as shown in Figure 37.16.
Fig. 37.16. (A) structure of immunoglobulin gene and the steps involved in DNA rearrangements in different regions. (B) a model explaining DNA rearrangements in heavy chain switching.
Enhancers in
regulation of immunoglobulin genes. Enhancers are certain DNA sequences, that increase the transcription of genes, although this is not the only characteristic of DNA segment to be called enhancer. Other essential characteristics of an enhancer are : (i) it is essential for transcription; (ii) it can be moved around on either side of gene or may be inverted and (iii) it can work with genes from sources other than its own. These enhancers have been discovered in immunoglobulin genes and have been found to work only in cells that make immunoglobulins. In this manner, enhancer is a remarkable regulatory sequence that is cell specific.
The enhancer sequence is found (i) between V-D-J and C in DNA segment for light chain and (ii) between V-J and C in DNA segment for heavy chain (Fig. 37.16). The role of enhancer sequence is demonstrated by the following evidences, (i) If intron region between V-D-J and C regions of heavy chain is removed from a cloned gene system, and then the latter is injected into mouse myeloma cells, the cloned genes lose their ability to make a heavy chain, (ii) When cloned intron region was cleaved into three fragments, and each is separately linked with genes for T-antigen, SV40, or β globin gene, only one of these fragments, presumably carrying the enhancer, stimulated the transcription, (iii) When cloned intron region of heavy chain gene was attached to
a bacterial gene, increased production of gene
product was achieved.
To obtain correct initiation of transcription, the enhancer must interact with other control sequences which are located upstream (towards 5' end) from the beginning of V-region. Normally V-D-J (or V-J joining the upstream sequence) and enhancer are far apart, but rearrangements bring them close enough for transcription to occur. The V-D-J segment may first attach to C
μ (which is first exon in C-region) and give I
gM. Later the cell may switch and may produce another immunoglobulin. This class switch may require further deletions of C-region exons. The deletion is mediated by a repeated DNA segment located just before each C-exon and designated as S-region (switch). The enhancer will function even after every class switch and thus can continue to activate transcription.




