Repression of transcription
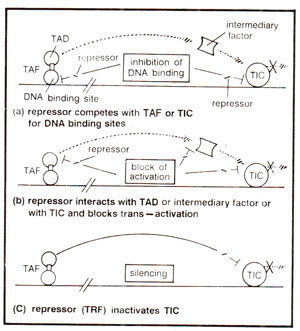
Fig. 37.5. Three categories of transcriptional repression (TAF = trans-activation factor; TIC = transcription initiation complex; TRF = trans-repressing factor).
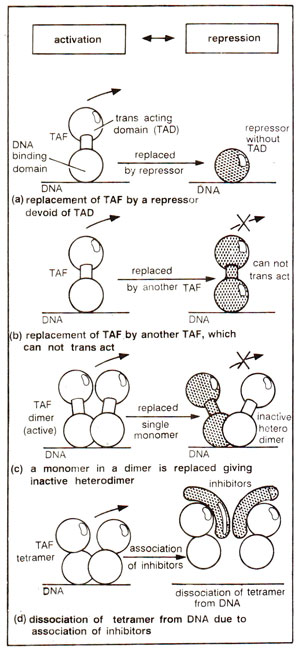
Fig. 37.6. Four ways in which DNA binding of TAF can be inhibited.
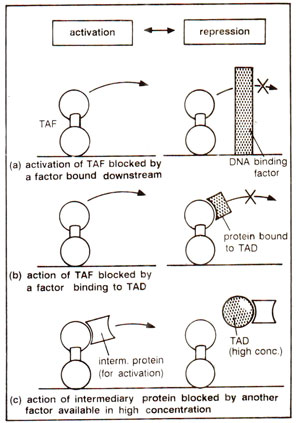
Fig. 37.7. Three ways in which activation (not binding on DNA) by TAF is blocked.
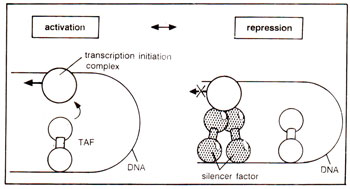
Fig. 37.8. Mehcanism of the action of silencer factors, rendering the transcription initiation complex ineffective.
Fig. 37.5. Three categories of transcriptional repression (TAF = trans-activation factor; TIC = transcription initiation complex; TRF = trans-repressing factor).
Inhibition of the binding of trans-activating factors (TAF) to DNA. One of the first examples of inhibition of TAF binding involved SV40 T antigen binding to its own promoter as in prokaryotes. In many other cases, there is a competition between a negative factor and a positive TAF as shown in the following examples : (i) In sea urchin, sperm specific histone gene H2B-1 is repressed in embryos, due to interference by a negative factor with a positive CCAAT binding factor, (ii) In humans, fetal γ-globin gene is repressed in adults due to binding of a factor NF-E, which inhibits binding of positive factor CP1 to CCAAT box. (iii) Several genes are repressed, when GC-box factor competes with Spl for binding to GC-rich sequences. During such an inhibition, a positive factor for one gene may also function as negative factor for another gene due to similar DNA sequences. This has been shown in several developmental genes in Drosophila and also for glucocorticoid receptor molecule, (iv) In still other cases, a component (e.g. Fos) of a heterodimer (e.g. Jun-Fos) can be replaced to give inactive complex (e.g. C-Jun-Jun B) which competes with heterodimers, or an inhibitor (e.g. IkB) may cause a TAF dessociate from DNA and inactivate it (Fig. 37.6). Heat shock protein (hsp 90) also associates with glucocorticoid receptor and renders it incapable of binding DNA.
Fig. 37.6. Four ways in which DNA binding of TAF can be inhibited.
Blocking of activation. In several examples, TAF can bind DNA, but its activation domain is not allowed to function (Fig. 37.7). Following alternatives are possible : (i) Negative factor may bind near the binding sites meant for positive factors. For instance, in c-myc gene, the negative factor myc-PRF binds next to positive factor myc-CFl, and for polyoma virus, repressor PEA2 binds near PEAl. (ii) Negative factor may bind the positive factor, rendering its trans-activation domain ineffective as done by binding of GAL-80 to TAF GAL4 in yeast;
Fig. 37.7. Three ways in which activation (not binding on DNA) by TAF is blocked.
Fig. 37.8. Mehcanism of the action of silencer factors, rendering the transcription initiation complex ineffective.
Support our developers




