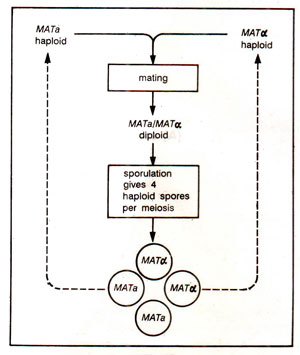
Fig. 37.17. Life cycle of yeast involving mating of MATa and MATαhaploids.
Life cycle of budding yeast,
S. cerevisiae (Fig. 37.17) shows that there are two mating types
(MATa and MATα), which fuse to give a diploid (MATa/MATα). The diploid cell sporulates and through meiosis forms four spores, two of each mating type. Each mating type is characterized by secretion of a
pheromone (MATa secretes
a factor and MATα secretes
a factor) and a
surface receptor for the pheromone of the opposite type. The diploids (MATa/MATα) produce no pheromone or surface receptor. The pheromone-receptor interaction activates a cell membrane protein called
G protein consisting of three subunits (α β γ). The activation needs a GTP molecule, which displaces GDP associated with αβγ
complex and releases αfrom βγ
dimer.
This separation of
a from βγ activates the next protein in signal transduction pathway. Therefore G protein is also called a
transducin. The dimer βγ alone (and not α) is capable of bringing about the activation. The above information has come from a study of mutations (called
STE =
sterile), which are incapable of mating. Atleast ten such mutations
(STE 1-7, 11, 12, 18) are known.
Fig. 37.17. Life cycle of yeast involving mating of MATa and MATαhaploids.
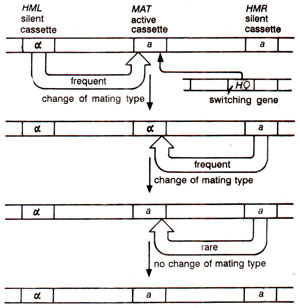
Fig. 37.18. Change of mating type in yeast due to replacement by active cassette by silent cassette of opposite type : when replacement involves cassettes of same type, the mating type remains unaltered.
A remarkable feature in yeast is the ability of some strains (which carry dominant
HO allele and not those which carry recessive
ho allele) to switch their mating type from
a to αand
vice versa. Consequently an
HO haploid strain, irrespective of its original mating type, will soon have cells of both mating types giving diploids (MATa/MATα). This is achieved by the presence of a
MAT (a or α) locus with an
active cassette (type
a or α) associated with two
silent cassettes at loci
HML(α
) on the left and
HMR(a) on the right. Only the active cassette at
MAT locus is expressed which can be
MATa or
MATα
. This can be replaced by
HMLαor
HMRa respectively leading to change in mating type (a→α or α→a).
MATa
is almost always replaced by,
HML and MATα by
HMR, so that if
HML is mutated to
HML a or
HMR is mutated to
HMRα
, replacement by them does not change mating type (Fig. 37.18). The
HML and
HMR are constitutively repressed (rendered silent) by some repressors binding to silencer sequences,
HML-E or
EL and
HMR-E or
ER located ~ 1 kb upstream of these loci (as discussed earlier). The switching is achieved by the activity of an endonuclease coded by thedominant gene
HO.
Fig. 37.18. Change of mating type in yeast due to replacement by active cassette by silent cassette of opposite type : when replacement involves cassettes of same type, the mating type remains unaltered.




