Single gene control of sex
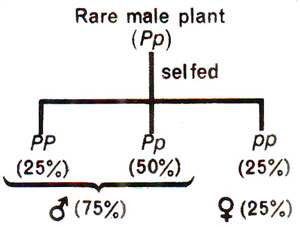
Fig. 17.21. Segregation for sex in seed obtained from a rare bisexual flower in Asparagus showing monogenic control.
Asparagus is a dioecious form. However, rarely female flowers bear rudimentary anthers and male flowers bear rudimentary pistils. Thus, rare male flowers having poorly developed pistils may set seeds. In one such case, when seeds obtained from a rare male flower, were raised into plants, male and female plants were found to be present in 3 : 1 ratio. When male plants raised thus were used to pollinate female flowers on female plants, only two-third of them showed segregation indicating that sex is controlled by a single gene. In this case, maleness should be dominant over femaleness and male plants should ordinarily be heterozygous. These results are diagrammatically represented in Figure 17.21.
Fig. 17.21. Segregation for sex in seed obtained from a rare bisexual flower in Asparagus showing monogenic control.
Maize is monoecious with male inflorescence (tassel) and female inflorescence (silk) located on the same plant. A gene tassel seed (ts) is known, which will convert the tassel into seed bearing inflorescence. Another gene silkless (sk) is responsible for the absence of silks. Therefore, a plant sk/sk will be effectively a male plant and a plant, ts/ts will be effectively a female plant. By using these genes it has been possible to convert maize from a monoecious to a dioecious form, where plants will bear only male or only female inflorescences.
For instance, if we use females with genotype ts ts sk sk (seeds on tassels, silkless) and males with genotype Ts ts sk sk (normal tassel and silkless) on crossing, they will give males and females (having same genotypes as parents) in 1 : 1 ratio. This will be stable dioecious situation.
In Drosophila, a transformer gene (tra) has been identified, which if present in homozygous condition (tra/tra) converts a female into a sterile male, but does not act upon normal male individual.
Support our developers




