Family Leguminosae
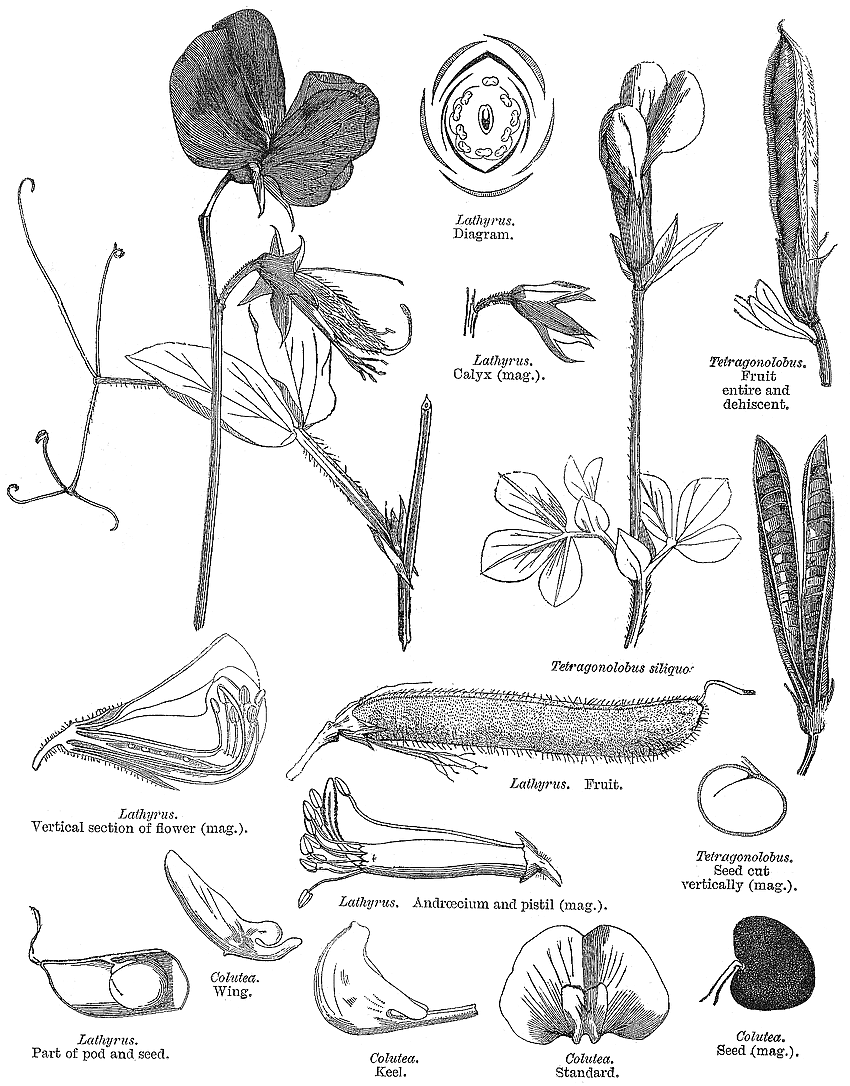
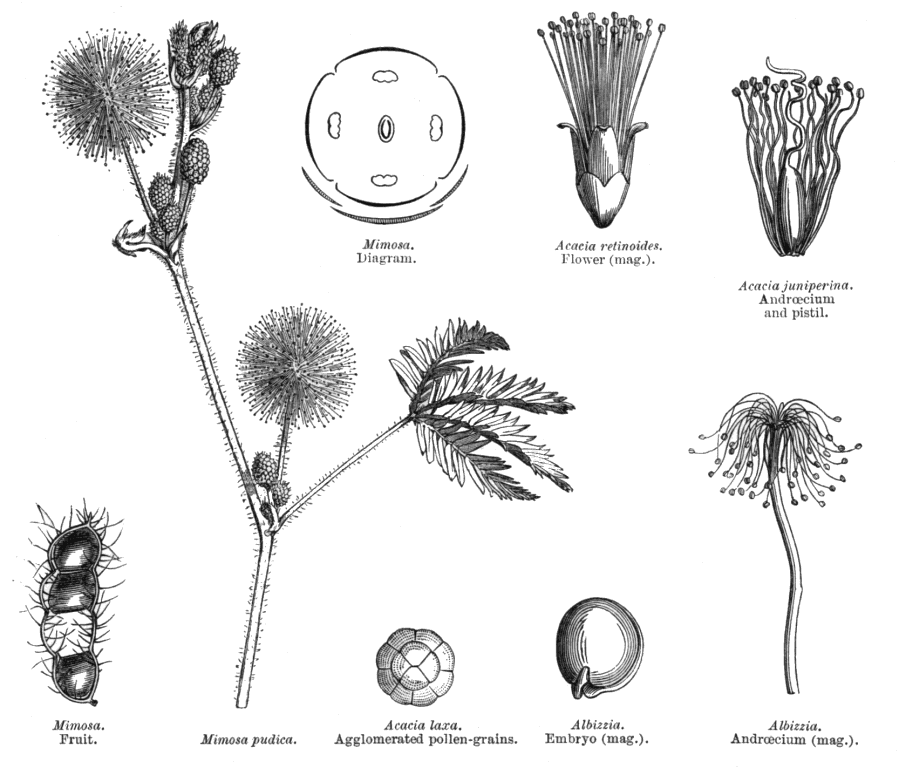
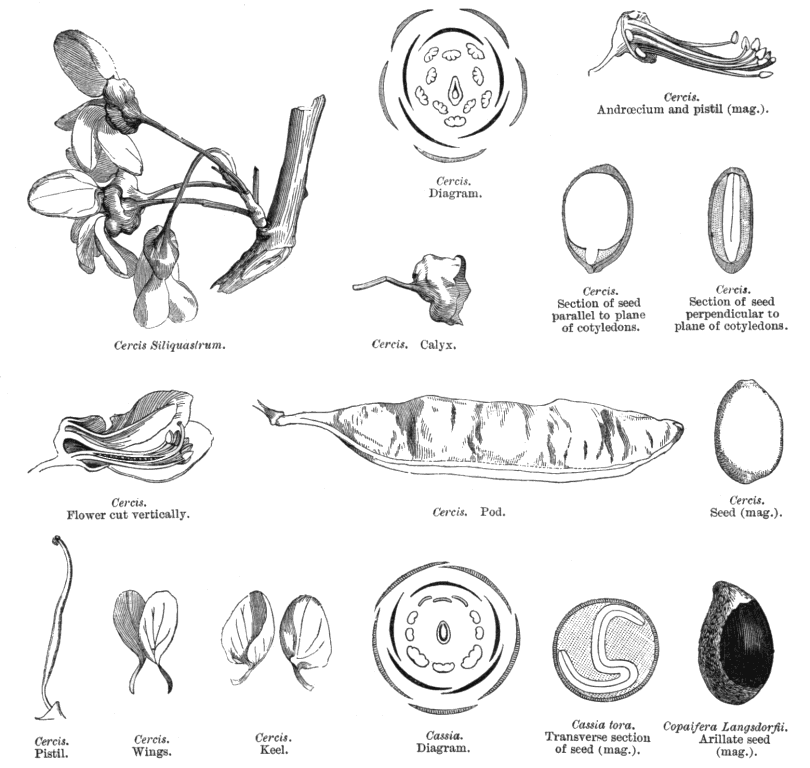
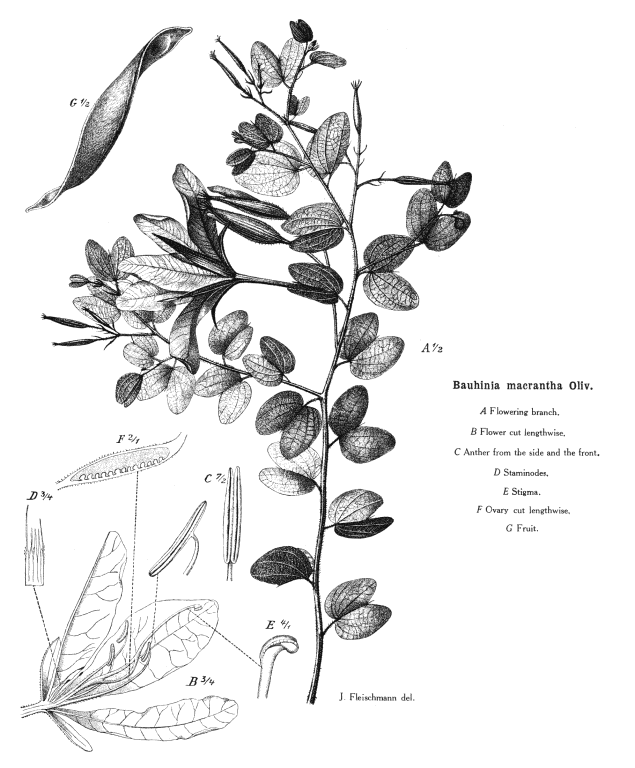
Leguminosae Juss.Alternatively Fabaceae Lindl. (but ambiguous) Including Caesalpiniaceae R.Br., Cassiaceae Link, Ceratoniaceae Link, , Detariaceae (DC.) Hess, Hedysareae (Hedysaraceae) J.G Agardh, Lathyraceae Burnett, Lotaceae Burnett, Mimosaceae R. Br., Papilionaceae Giseke, Phaseolaceae Ponce de Léon & Alvares, Robiniaceas (Robiniaceae) Welw., Swartzieae (Swartziaceae (DC.) Bartl. Habit and leaf form. Trees, or shrubs, or herbs, or lianas; resinous, or not resinous. ‘Normal’ plants, or switch-plants; the switch forms often with the principal photosynthesizing function transferred to stems, or phyllodineous. Leaves well developed (usually), or much reduced (not infrequently). The herbs annual, or biennial, or perennial; with neither basal nor terminal aggregations of leaves. Self supporting, or epiphytic, or climbing; the climbers stem twiners, or tendril climbers (via stem or leaf tendrils), or scrambling (then sometimes via hooks); the twiners twining clockwise, or twining anticlockwise (in Phaseolus, Wisteria). Helophytic, or mesophytic, or xerophytic. Heterophyllous (e.g. Acacias with bipinnate juvenile and phyllodineous mature foliage), or not heterophyllous. Leaves evergreen, or deciduous; minute to very large; alternate (usually), or opposite to whorled (e.g. some Mirbelieae); spiral, or distichous; ‘herbaceous’, or leathery, or membranous, or modified into spines; petiolate to sessile; non-sheathing; gland-dotted, or not gland-dotted; aromatic, or without marked odour (usually); edgewise to the stem (commonly when phyllodineous, especially in Australia), or with ‘normal’ orientation; compound (commonly), or simple; pulvinate, or epulvinate; when compound (as is usual) unifoliolate, or ternate, or pinnate (commonly, either pari- or imparipinnate), or palmate, or bipinnate (commonly), or bifoliolate (e.g. Bauhinieae). Leaflets pulvinate, or epulvinate. Leaves stipulate (nearly always), or exstipulate (e.g., some Mirbelieae). Stipules intrapetiolar; scaly, or leafy, or spiny, or represented by glands; caducous, or persistent. Leaves without a persistent basal meristem. Leaf anatomy. Mucilaginous epidermis present, or absent. Stomata anomocytic, or paracytic, or anisocytic, or tetracytic, or cyclocytic. Urticating hairs absent (but present on calyces and pods of Mucuna). Lamina dorsiventral, or isobilateral, or centric; with secretory cavities, or without secretory cavities. Secretory cavities containing oil, or containing mucilage, or containing resin. The mesophyll containing mucilage cells, or not containing mucilage cells; with sclerencymatous idioblasts (occasionally), or without sclerenchymatous idioblasts. Minor leaf veins with phloem transfer cells (55 genera, including some from each subfamily), or without phloem transfer cells (67 genera. Watson and Gunning (1981) detail Caesalpinioideae and Papilionoideae. For Mimosoideae, Pate and Gunning (1969) recorded as positive species of Mimosa, Neptunia and (dubiously) Calliandra; they recorded negatives for Acacia, Albizzia, Adenenanthera, Dichrostachys, Enterolobium, Leucaena, Pithecellobium, Prosopis and Wallaceodendron). Stem anatomy. Secretory cavities present, or absent. Cork cambium present (usually), or absent; initially deep-seated, or superficial. Nodes tri-lacunar, or penta-lacunar. Primary vascular tissue in a cylinder, without separate bundles, or comprising a ring of bundles. Cortical bundles present, or absent. Medullary bundles absent. Internal phloem absent. Secondary thickening absent, or developing from a conventional cambial ring, or anomalous; when anomalous, via concentric cambia (e.g. Koompassia, Derris, Mucuna, Wisteria). The secondary phloem stratified into hard (fibrous) and soft (parenchymatous) zones, or not stratified. ‘Included’ phloem present, or absent. Xylem with vessels. Vessel end-walls simple. Vessels with vestured pits, or without vestured pits. Wood storied, or partially storied (VPI); parenchyma apotracheal, or paratracheal. Sieve-tube plastids P-type (mostly), or S-type (in a few Papilionoideae only); when P-type type IV (subtype (a) in Mimosoideae, (b) elsewhere). Reproductive type, pollination. Plants hermaphrodite (mostly, in Caesalpinioideae and Papilionoideae), or monoecious, or andromonoecious, or polygamomonoecious (unisexual flowers commoner in Mimosoideae). Pollination entomophilous, or ornithophilous (especially common in southern Australia), or cheiropterophilous (e.g. Mucuna holtoni, where the nectar guide is a petal functioning as a ‘concave mirror’ for ultrasound); mechanism conspicuously specialized (with at least two forms of passive presenter, involving modifications of the style and/or of the keel of the corolla and/or the staminal filaments, and explosive pollination in (e.g.) Medicago), or unspecialized. Inflorescence, floral, fruit and seed morphology. Flowers aggregated in ‘inflorescences’ (usually), or solitary; in panicles, in fascicles, in racemes, in spikes, and in heads. The ultimate inflorescence unit cymose, or racemose (— often ostensibly racemose, but frequently paniculate or, as in many Phaseoleae, having pseudoracemes bearing nodal clusters of obscure constitution). Inflorescences terminal, or axillary, or leaf-opposed (e.g. in some Bossiaeeae); pseudanthial (e.g. Mimosoideae), or not pseudanthial. Flowers minute to large; regular (Mimosoideae), or somewhat irregular to very irregular (Papilionoideae, most Caesalpinioideae); commonly zygomorphic; resupinate (sometimes, in association with bird pollination or in pendulous inflorescences), or not resupinate. The floral irregularity involving the perianth and involving the androecium. Flowers papilionaceous (Papilionoideae), or ‘pseudo-papilionaceous’ (‘ascending’ in most Caesalpinioideae), or neither papilionaceous nor pseudo-papilionaceous (regular in Mimosoideae); tetracyclic (mostly), or pentacyclic to polycyclic. Floral receptacle developing a gynophore (this often adnate to the hypanthium in Caesalpinioideae), or with neither androphore nor gynophore; usually more or less cupular. Free hypanthium present, or absent (more or less replaced by the calyx tube in most Papilionoideae). Hypogynous disk present, or absent. Perianth with distinct calyx and corolla (nearly always), or sepaline (corolla at least sometimes absent in 26 genera of Caesalpinioideae, some Swartzieae, some Amorphieae); (3–)5, or (6–)10(–11); 1 whorled (rarely), or 2 whorled; isomerous, or anisomerous. Calyx 5, or (3–)5(–6); 1 whorled; polysepalous, or partially gamosepalous, or gamosepalous (characteristic of most Mimosoideae and Papilionoideae). In Caesalpinioideae, sometimes 2 of the members joined. Calyx unequal but not bilabiate, or bilabiate, or regular; persistent (usually), or not persistent (e.g., Lamprolobium); accrescent (rarely), or non-accrescent; imbricate, or valvate (or splitting irregularly in the Swartzieae); when pentamerous, with the median member anterior. Epicalyx present (e.g., Pultenaea), or absent (usually). Corolla (1–)3–5 (commonly reduced below five or missing in Swartzieae, Amorphieae and Caesalpinioideae, commonly 3–4 in Mimosoideae); 1 whorled; appendiculate (petals variously auriculate, lobed, etc.), or not appendiculate; polypetalous (commonly in Caesalpinioideae), or partially gamopetalous (usually, in Papilionoideae), or gamopetalous (in some Mimosoideae and Sympetalandra). Papilionoideae commonly with 2 of the petals joined (the two ventral petals connivent to form the corolla ‘keel’), or 4 of the petals joined (the wings adnate to the keel). The joined petals of the papilionate corolla anterior (or anterior and lateral). Corolla imbricate (descending in Papilionoideae, ascending in Caesalpinioideae), or valvate (Mimosoideae), or with open aestivation (occasionally); white, or yellow, or orange, or red, or pink, or purple, or blue; or some members persistent (e.g. Trifolium), or deciduous. Petals clawed, or sessile. Androecium (1–)9–10, or 10–50 (often 9–10, but commonly fewer, especially in Caesalpinioideae, and sometimes ‘many’ in Mimosoideae, Swartzieae, Sophoreae). Androecial members free of the perianth (mostly), or adnate (e.g. in Caesalpinioideae-Parkieae, Dalbergieae, Mirbelieae, Trifolium, Genista, etc., where at least some members or the androecial tube can be attached to corolla components); all equal, or markedly unequal; free of one another (sometimes), or coherent (in a variety of configurations); when cohering 1 adelphous, or 2 adelphous (commonly with the tenth, posterior stamen free of the rest, whose filaments are united into a tube); 1 whorled (when five, nine or ten, though the antesepalous, theoretically ‘outer’ members develop first, are often longer, and their anthers may differ from those of the antepetalous members), or 2–6 whorled (? in some Mimosoideae). Androecium exclusively of fertile stamens (usually, and nearly always in Papilionoideae), or including staminodes. Stamens (1–)9–10(–50); reduced in number relative to the adjacent perianth, or isomerous with the perianth, or diplostemonous to polystemonous. Anthers separate from one another, or connivent; dorsifixed, or basifixed, or dorsifixed and basifixed (alternating); versatile (commonly), or non-versatile; dehiscing via pores, or dehiscing via longitudinal slits; introrse, or latrorse; bilocular (usually), or unilocular to bilocular (the thecae sometimes confluent above); tetrasporangiate; appendaged, or unappendaged. Endothecium developing fibrous thickenings. Microsporogenesis simultaneous. Anther wall initially with one middle layer, or initially with more than one middle layer; of the ‘dicot’ type. Tapetum usually glandular. Pollen shed in aggregates (often in Mimosoideae, infrequently elsewhere, e.g. Afzelia), or shed as single grains (usually); when aggregated, in tetrads, or in polyads. Pollen grains aperturate (usually), or nonaperturate; (2–)3(–4) aperturate, or 6 aperturate; colporate (commonly), or porate (the pores sometimes operculate), or colpate, or rugate; 2-celled (nearly always), or 3-celled (rarely, in Mimosoideae). Gynoecium 1 carpelled (nearly always), or 2–16 carpelled (in a few Mimosoideae from New Guinea and tropical Australia (Archidendron) and tropical South America (Affonsea, Klugiodendron); partly petaloid (Petalostylis), or non-petaloid. Carpels reduced in number relative to the perianth (nearly always), or isomerous with the perianth to increased in number relative to the perianth (rarely). The pistil 1 celled (nearly always), or 2 celled (by a false septum, e.g. Mirbelia). Gynoecium monomerous (usually), or apocarpous; of one carpel (usually), or eu-apocarpous (rarely); superior. Carpel apically stigmatic; (1–)2–100 ovuled (i.e. to ‘many’, usually in alternating rows along the placenta). Placentation marginal (along the ventral suture). Gynoecium median (the placenta posterior, on the ventral suture). Ovary sessile to stipitate. Ovules pendulous to ascending; biseriate; arillate, or non-arillate; anatropous, or campylotropous to amphitropous, or hemianatropous; bitegmic; crassinucellate. Outer integument contributing to the micropyle, or not contributing to the micropyle. Embryo-sac development Polygonum-type. Polar nuclei fusing prior to fertilization. Antipodal cells formed; 3; not proliferating; ephemeral (Papilionoideae, Mimosoideae), or persistent (most Caesalpinioideae). Synergids hooked (often with filiform apparatus). Endosperm formation nuclear. Endosperm haustoria present; chalazal, or lateral (rarely). Embryogeny onagrad, or asterad, or caryophyllad. Fruit non-fleshy, or fleshy. The fruiting carpel dehiscent, or indehiscent; a legume (usually), or a follicle, or an achene, or samaroid, or a loment, or drupaceous. Fruit elastically dehiscent, or passively dehiscent. Gynoecia of adjoining flowers combining to form a multiple fruit (rarely, in Mimosoideae), or not forming a multiple fruit. Dispersal unit the seed, or the fruit. Fruit (1–)2–100 seeded (to ‘many’). Seeds endospermic, or non-endospermic (endosperm copious only in some Caesalpinioideae); small to very large; winged (e.g. in some Mimosoideae), or wingless (usually). Seeds with starch, or without starch. Seeds with amyloid, or without amyloid. Cotyledons 2; usually flat. Embryo chlorophyllous (45/83 — representing all three subfamilies); straight, or curved, or bent (the radicle straight in Mimosoideae, straight or oblique in Caesalpinioideae, usually inflexed in Papilionoideae). The radicle when flexed, lateral. Micropyle zigzag, or not zigzag. Seedling. Germination phanerocotylar, or cryptocotylar. Physiology, biochemistry. Nitrogen-fixing root nodules present (seemingly the norm in Papilionoideae and Mimosoideae), or absent (seemingly, from many Caesalpinioideae). Cyanogenic (rarely), or not cyanogenic (mostly). Cynogenic constituents tyrosine-derived, or phenylalanine-derived, or of Hegnauer’s ‘Group C’, or leucine-derived. Alkaloids present (commonly), or absent. Iridoids not detected. Proanthocyanidins present, or absent; when present, cyanidin, or delphinidin, or cyanidin and delphinidin. Flavonols present (mostly), or absent; kaempferol and quercetin, or quercetin and myricetin, or kaempferol, quercetin, and myricetin. Ellagic acid consistently absent (from 54 species and 41 genera, representing all three subfamilies). Arbutin present, or absent. Aluminium accumulation not found. Sugars transported as sucrose (in numerous species and genera from all three subfamilies). C3. C3 physiology recorded directly in Acacia, Alysicarpus, Amorpha, Arachis, Astragalus, Caragana, Cassia, Cercidium, Crotalaria, Dalea, Dolichos, Genista, Gleditsea, Glycine, Hoffmanseggia, Indigofera, Lespedeza, Lotus, Lupinus, Medicago, Mimosa, Olneya, Phaseolus, Pisum, Prosopis, Pueraria, Robinia, Sesbania, Spartium, Stylosanthes, Tephrosia, Trifolium, Vicia, Vigna. Anatomy non-C4 type (recorded from numerous genera representing all three sybfamilies). Geography, cytology. Holarctic, Paleotropical, Neotropical, Cape, Australian, and Antarctic. Frigid zone, temperate, sub-tropical, and tropical. Cosmopolitan. Taxonomy. Subclass Dicotyledonae; Crassinucelli. Dahlgren’s Superorder Fabiflorae; Fabales. Cronquist’s Subclass Rosidae; Fabales. APG 3 core angiosperms; core eudicot; Superorder Rosanae; fabid; Order Fabales. Species 12000. Genera about 650; Mimosoideae: Albizia, Inga, Pithecellobium, Acacia, Mimosa, Prosopis, Piptadenia, Entada, Parkia, etc. Caesalpinioideae: Caesalpinia, Parkinsonia, Delonix, Gleditsea, Haematoxylum, Sclerolobium, Melanoxylon, Bauhinia, Cercis, Cassia, Ceratonia, Colophospermum, Copaifera, Brachystegia, Tamarindus, Amherstia, etc. Papilionoideae: Sophora, Baphia, Podalyria, Glycine, Phaseolus, Vigna, Crotalaria, Lupinus, Cytisus, Medicago, Trifolium, Lotus, Indigofera, Astragalus, Vicia, Lathyrus, Pisum, Arachis, Aeschynomene, etc. Clearly, the many features which tend distinguish the subfamilies all involve rather numerous exceptions, are very incompletely documented, or are not universally applicable. Economic uses, etc. Economically very important for food, fodder, fibres, dyes, gums, resins, oils, and ‘green manure’; e.g. peas (Pisum), lentils (Lens), peanuts (Arachis), beans (Phaseolus, Vicia), cowpeas (Vigna), soybean (Glycine), clover (Trifolium), alfalfa (lucerne, Medicago), lupins (Lupinus), sweet clover (Melilotus). Numerous cultivated ornamentals, e.g. Bauhinia, Wisteria, Acacia, Cassia, Cytissus, Genista, Albizia, Lathyrus. Important tropical timbers from Acacia, Albizzia, Dalbergia, Robinia, Sophora, etc. Illustrations. Quotations This fellow pecks up wit as pigeons peas . . . . luscious as locusts Like a richly coloured map For Linnaeus |
Support our developers





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}