Family Ceratophyllaceae
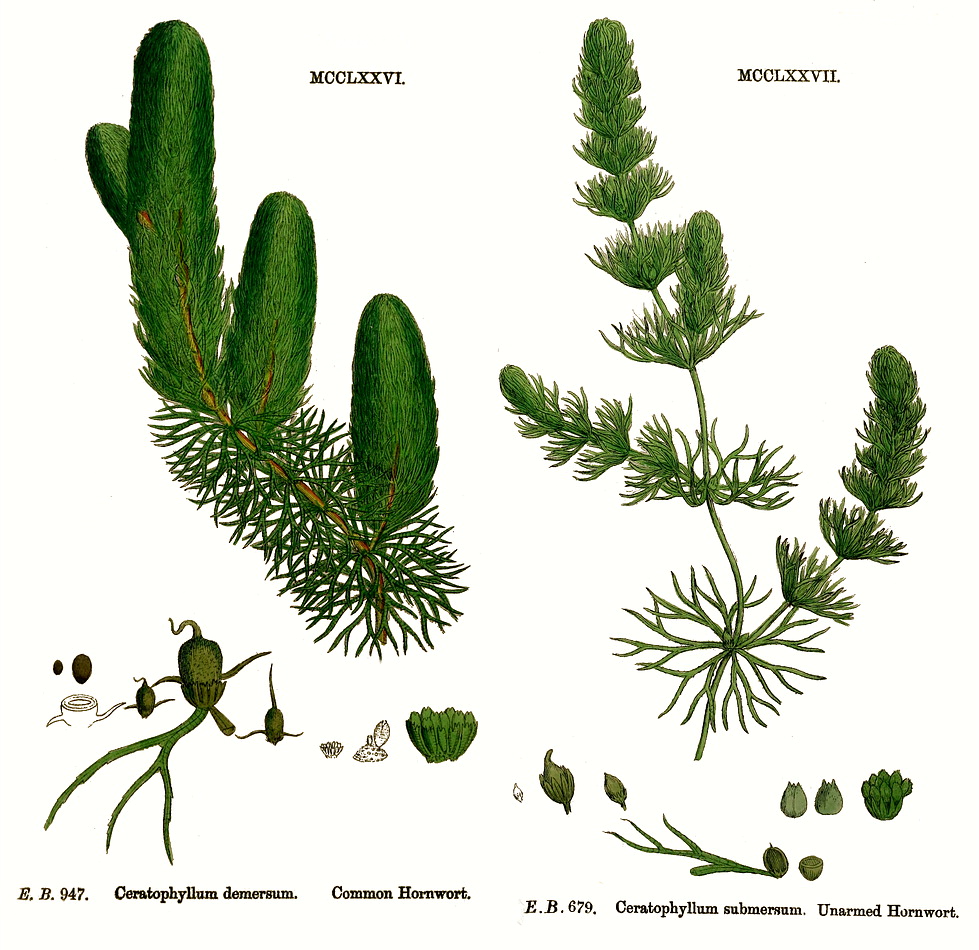
Ceratophyllaceae S.F. GrayHabit and leaf form. Submerged, brittle, rootless aquatic herbs (the plant decaying behind as it grows in front). Plants rootless. With neither basal nor terminal aggregations of leaves. Hydrophytic; free floating. Leaves submerged. Leaves medium-sized; whorled; 3–10 per whorl; becoming brittle with age; petiolate; simple. Lamina dissected; finely dichotomously dissected. Leaves exstipulate. Leaf anatomy. Minor leaf veins without phloem transfer cells. Stem anatomy. Secondary thickening absent. Xylem without vessels. Reproductive type, pollination. Fertile flowers functionally male and functionally female. Plants monoecious. Female flowers without staminodes. Pollination by water. Inflorescence, floral, fruit and seed morphology. Flowers solitary, or aggregated in ‘inflorescences’ (one per leaf axil, in whorls!); axillary; ebracteate; ebracteolate; minute; regular; cyclic (female), or partially acyclic (male). The androecium acyclic. Hypogynous disk absent. Perianth sepaline (bractlike); 9–10; joined (basally); 1 whorled. Androecium (5–)10–20(–27) (in male flowers). Androecial members free of the perianth; all equal; free of one another. Androecium exclusively of fertile stamens. Stamens (5–)10–20(–27); filantherous to with sessile anthers (‘subsessile’). Anthers adnate; non-versatile; dehiscing irregularly, or dehiscing via longitudinal slits; extrorse; appendaged (via the often coloured, apically projecting connective). The anther appendages apical. Microsporogenesis successive, or simultaneous. Tapetum amoeboid, or glandular. Pollen shed as single grains. Pollen grains nonaperturate; 2-celled. Gynoecium in female flowers 1 carpelled. The pistil 1 celled. Gynoecium monomerous; of one carpel; superior. Carpel stylate; apically stigmatic; 1 ovuled. Placentation apical. Ovules pendulous; non-arillate; orthotropous; unitegmic; crassinucellate. Embryo-sac development Polygonum-type. Polar nuclei fusing prior to fertilization. Antipodal cells formed; 3; not proliferating; small. Hypostase present. Endosperm formation cellular. Embryogeny asterad. Fruit non-fleshy. The fruiting carpel indehiscent; an achene. Seeds non-endospermic. Embryo well differentiated (but without a functional radicle). Cotyledons 2. Embryo chlorophyllous (fleshy); straight. Seedling. Germination phanerocotylar. Physiology, biochemistry. Not cyanogenic. Proanthocyanidins present; cyanidin and delphinidin. Flavonols absent. Ellagic acid absent (C. demersum). Aluminium accumulation not found. Geography, cytology. Temperate to tropical. Cosmopolitan. X = 12. Taxonomy. Subclass Dicotyledonae; Crassinucelli. Dahlgren’s Superorder Nymphaeiflorae; Nymphaeales. Cronquist’s Subclass Magnoliidae; Nymphaeales. APG 3 core angiosperms; peripheral eudicot (but not overtly assigned as such); Superorder Ceratophyllanae; Order Ceratophyllales. Species 10. Genera 1; only genus, Ceratophyllum. See Les (1993), who suggested that Ceratophyllum may have arisen from angiosperms pre-dating the divergence of Monocots and Dicots. It has subsequently been proposed (Chase et al., 1993) as ‘sister to all other flowering plants’, consequent on ‘phylogenetic analyses’ of RuBisCO large subunit DNA sequences. It should be remembered, however, that RuBisCOs of all the submerged aquatics assayed for KmCO2 (Yeoh et al. 1981: sample including algae, bryophytes, Monocots, and both tenuinucellate and crassinucellate Dicots, but not Ceratophyllum) exhibit peculiarly high values, implying peculiar nucleotide sequences in the RuBisCO genes. Illustrations. |
Support our developers





{kind=link}
{kind=link}
{kind=link}