Chloroplasts
The photosynthetic compartment contains the pigments for absorbing light and channeling the
energy of the excited pigment molecules into a series of photochemical and enzymatic reactions.
These pigments are organized in proteic complexes embedded in the membrane of sac-like flat
compressed vesicles known as the thylakoids. These vesicles are about 24 nm thick, and enclose
a space, termed lumen, 10 nm wide.
In prokaryotes the thylakoids are free within the cytoplasm, while in eukaryotes they are
enclosed within bounding membranes to form the chloroplast. The colorless matrix of the chloroplast
is known as the stroma. Inside the chloroplast, thylakoids are organized into two different
compartments, granal thylakoids, stacked into hollow disks termed grana; and stromal thylakoids
forming multiple connections between the grana. All thylakoids surfaces run parallel to the
plane of the maximum chloroplast cross-section.
Chloroplasts contain nucleic acids and ribosomes. DNA is naked, that is, not associated with proteins, and occurs in two configurations: scattered, but not connected, small nucleoids or as a peripheral ring. Chloroplasts are semiautonomous organelles that replicate their own DNA, and this replication is not linked to the division of the organelle. Synthesis of RNA and proteins is possible inside the chloroplast, though they are not strictly autonomous from the nuclear genome. The plastid genes are transcribed and translated within the plastids. The machinery of protein synthesis is, in any case, partially composed of imported nuclear gene products. In this respect, as in others, the chloroplast is no longer independent. Nevertheless, about half of the plastid genome consists of genes that contribute to the machine of gene expression, for example, genes for rRNA, tRNAs, RNA polymerase subunits, and ribosomal proteins. Plastids code for, and synthesize, some proteins that are components for photosystems and particular subunits of photosynthetic enzymes. The missing subunits of these complexes are coded in the nucleus and must be imported from the cytoplasm.
Chloroplast development and division (self-replication) may be coordinated with that of the cell
or may proceed independently. They divide in mother cells and they are inherited by daughter cells
during vegetative division and usually only from the maternal side in sexual reproduction. The
shape and number of chloroplasts are extremely variable from the single cup-shaped chloroplast
of Dunaliella salina, the ribbon-like chloroplast of
Spirogyra or stellate one of
Zygnema to the numerous (about 10
8) ellipsoidal chloroplasts of
Acetabularia giant cell.
The fact that algae of different divisions have different colors, due to the presence of a variety of pigments in the photosynthetic membrane system, might lead to the supposition that the photosynthetic membrane structures are variable. This is true because the features of photosynthetic membrane system represent a diagnostic important element at the class level. In this section of anatomy we will consider the structure, composition and location of the photosynthetic membrane system in each algal division.
Cyanophyta and ProchlorophytaThe photosynthetic apparatus of these algae is localized on intracytoplasmic membranes termed
thylakoids. The thylakoids membranes show considerable variations in structure and arrangements
depending on the species. Also the amount of thylakoid membranes per cell is variable due to
growth conditions and taxonomic specificity.
In most species the thylakoids are arranged peripherally in three to six layers running parallel to
the cell membrane, forming an anastomosing network of concentric shells. This peripheral region
has been called the chromatoplast, which is separated from the inner nucleoplasmic region called
the centroplasm. This type of thylakoid arrangement is characteristic of many unicellular and
filamentous cyanobacteria, such as
Synechococcus planctibus and
Anabaena sp.
In some other organisms such as
Oscillatoria and
Arthrospira the thylakoids are orientated perpendicular to the longitudinal cell wall. Radial arrangement is present in
Phormidium retzi. Thylakoids are not always restricted to the cell periphery but can be found scattered throughout the cell as in Gloeotrichia sp. However, the arrangement of thylakoids can change from cell to cell and cell types (vegetative cells, heterocyst, akinete) within the same culture from parallel to a convoluted appearance. Thylakoids can fuse with each other and form an anastomosing network; unlike the
chloroplasts of eukaryotic algae, the thylakoids of cyanobacteria form stacking regions only to a
very limited extent. During cell division the thylakoids have to be separated and divided into the
daughter cells; generally this division is an active process. Cell division starts by a centripetal
growth of cytoplasmic membrane and peptidoglycan layer, to form the septum; the thylakoid cylinder
is narrowed at the level of the cross wall by invagination before the septum is formed.
Proteins,
lipids, carotenoids, and chlorophyll a are major components of thylakoids. On the outer surface of
the thylakoids regular rows of electron dense granular structures are closely attached. These granules
termed phycobilisomes contain the light harvesting phycobiliproteins, that is, allophycocyanin,
phycocyanin, and phycoerythrin (Figure 2.75). The phycobilisome structure consists of a threecylinder
core of four stacked molecules of allophycocyanin, closest to the thylakoid membrane,
on which converge rod-shaped assemblies of coaxially stacked hexameric molecules of only phycocyanin
or both phycocyanin and phycoerythrin (Figure 2.76). Phycobiliproteins are accessory
pigments for the operation of photosystem II also in Glaucophyta, Cryptophyta, and Rhodophyta.

FIGURE 2.75 Transmission electron microscopy image of a cyanobacterium in longitudinal section, showing the thylakoid membranes with phycobilisomes. (Bar: 0.20 µm.) (Courtesy of Dr. Luisa Tomaselli.)
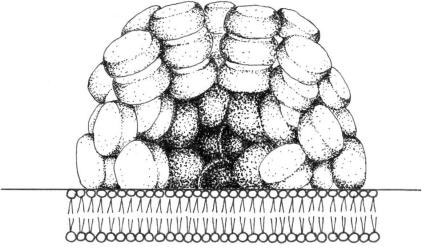
FIGURE 2.76 Schematic drawing of a phycobilisome.
The thylakoid membranes of Prochloron (Prochlorophyta) are organized into stacked and
unstacked regions reminiscent of the grana and stromal lamellae of higher plant chloroplasts.
They differ from cyanobacteria in that they contain chlorophyll a/b light harvesting systems
rather than phycobiliproteins organized in the phycobilisomes.




