Family Icacinaceae
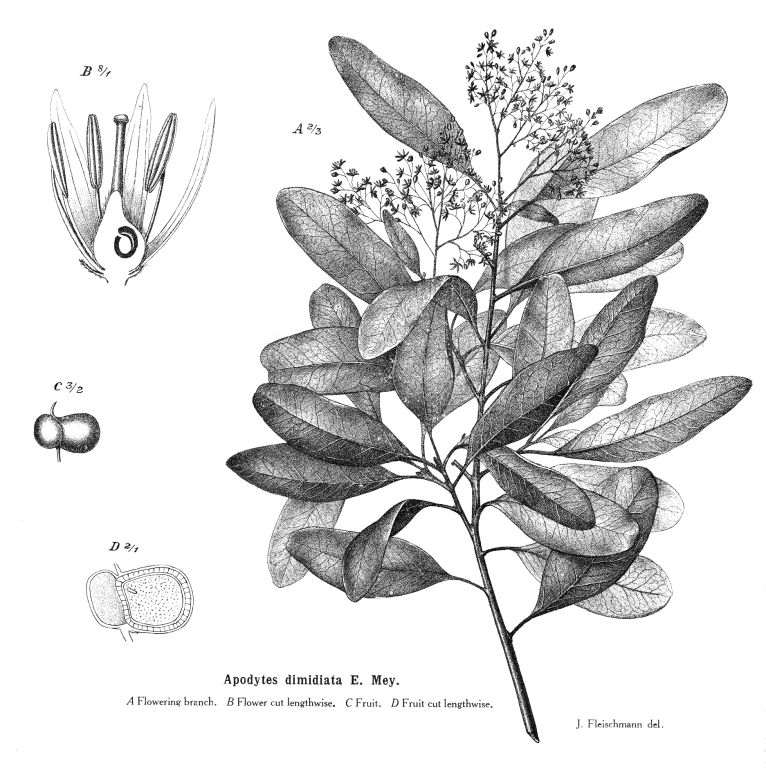
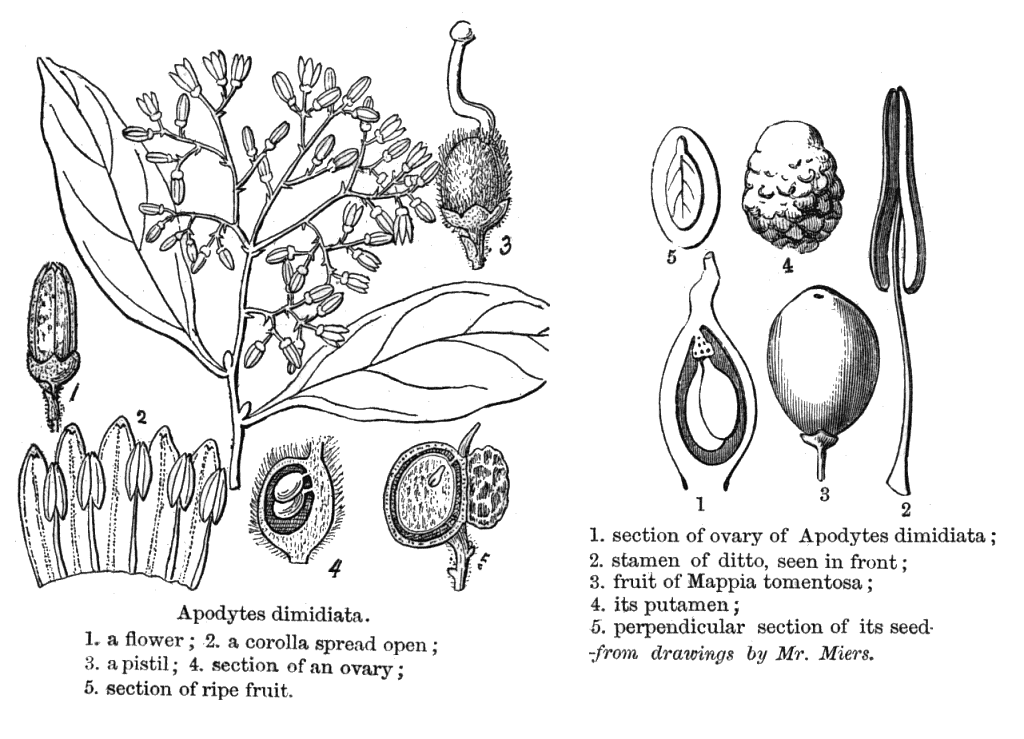
Icacinaceae MiersAlternatively Irvingbaileyaceae Including Emmotaceae Van Tiegh., Iodaceae Van Tiegh., Leptaulaceae Van Tiegh., Phytocreneae (Phytocrenaceae) R.Br., Pleurisanthaceae Van Tiegh., Sarcostigmataceae Van Tiegh., StemonuraceaeExcluding Pennantiaceae J.G. Agardh Habit and leaf form. Trees, shrubs, and lianas; laticiferous, or non-laticiferous and without coloured juice. Self supporting, or climbing. Helophytic, or mesophytic. Leaves alternate (nearly always), or opposite (Iodes); spiral; leathery; petiolate; non-sheathing; not gland-dotted; simple. Lamina entire; pinnately veined; cross-venulate. Leaves exstipulate. Lamina margins entire to dentate. Leaves without a persistent basal meristem. Domatia occurring in the family (4 genera); manifested as pits, or pockets. Leaf anatomy. Hydathodes present (occasionally), or absent. Mucilaginous epidermis present, or absent. Stomata anomocytic, or anisocytic, or paracytic. Lamina with secretory cavities (when present,), or without secretory cavities. Secretory cavities containing latex. The mesophyll with sclerencymatous idioblasts, or without sclerenchymatous idioblasts. Minor leaf veins without phloem transfer cells (Cassinopsis). Stem anatomy. Secretory cavities present, or absent; when present, with latex. Cork cambium present; initially superficial. The cortex containing cristarque cells, or without cristarque cells. Nodes unilacunar, or tri-lacunar. Primary vascular tissue often bicollateral. Medullary bundles present, or absent. Internal phloem absent. Secondary thickening developing from a conventional cambial ring, or anomalous; when anomalous, via concentric cambia, or from a single cambial ring. ‘Included’ phloem present (from a single cambial ring, in lianoids of the tribe Sarcostigmateae), or absent. Xylem with tracheids, or without tracheids; with fibre tracheids, or without fibre tracheids; with vessels. Vessel end-walls scalariform, or simple, or scalariform and simple. Vessels without vestured pits. Reproductive type, pollination. Plants hermaphrodite (usually), or andromonoecious, or gynomonoecious, or dioecious (especially the lianas), or androdioecious, or gynodioecious, or polygamomonoecious (‘rarely polygamous to dioecious’). Inflorescence, floral, fruit and seed morphology. Flowers aggregated in ‘inflorescences’ (the pedicels articulated); in cymes, in panicles, in racemes, and in spikes. The ultimate inflorescence unit cymose, or racemose. Inflorescences axillary (usually), or leaf-opposed, or epiphyllous (Leptaulus). Flowers small; odourless; regular; (3–)4–5(–6) merous; cyclic; tetracyclic. Free hypanthium absent. Hypogynous disk absent (usually), or present; when present, extrastaminal, or intrastaminal. Perianth with distinct calyx and corolla (usually), or sepaline (rarely apetalous); (3–)4–5(–6), or (6–)8–10(–12); 2 whorled; isomerous. Calyx when present (i.e. usually), (3–)4–5(–6); 1 whorled; gamosepalous (tubular); blunt-lobed; regular; non-accrescent; imbricate (usually), or valvate (rarely). Corolla when present (i.e. usually), (3–)4–5(–6); 1 whorled; polypetalous (usually), or gamopetalous (rarely); usually valvate; regular. Androecium (3–)4–5(–6). Androecial members free of the perianth, or adnate (to the corolla tube); all equal; free of one another; 1 whorled. Androecium exclusively of fertile stamens. Stamens (3–)4–5(–6); isomerous with the perianth; oppositisepalous; alternating with the corolla members. Anthers dehiscing via longitudinal slits; usually introrse; unappendaged. Endothecium developing fibrous thickenings. Microsporogenesis simultaneous. The initial microspore tetrads tetrahedral. Anther wall initially with one middle layer to initially with more than one middle layer (these ephemeral). Tapetum glandular. Pollen grains aperturate, or nonaperturate (Stachyanthus); when aperturate, mostly 3(–7) aperturate; colpate, or porate, or colporate, or foraminate; 3-celled (in Miquelia and Phytocrene). Gynoecium (2–)3(–5) carpelled. The pistil 1 celled (usually), or 3–5 celled. Gynoecium syncarpous; synstylovarious to eu-syncarpous; superior. Ovary 1 locular (usually, by abortion), or 3–5 locular (rarely). Gynoecium stylate (usually shortly). Styles 1; attenuate from the ovary; apical. Stigmas (2–)3(–5). Placentation usually apical; when plurilocular (i.e. rarely) axile, or apical. Ovules in the single cavity usually 2 (back to back — rarely only one); when plurilocular (i.e. rarely) 1–2 per locule; pendulous; arillate (with a funicular thickening near the micropyle), or non-arillate; anatropous; unitegmic; crassinucellate (e.g., Icacina, Iodes, Pyrenacantha, Stemonurus), or tenuinucellate (Phytocrene). Embryo-sac development Polygonum-type. Polar nuclei fusing prior to fertilization. Antipodal cells formed; 3; not proliferating; ephemeral, or persistent. Synergids hooked (with filiform apparatus). Endosperm formation nuclear. Endosperm haustoria present, or absent; when present, chalazal (Nothapodites). Fruit fleshy, or non-fleshy; indehiscent; a drupe (usually), or a samara (sometimes). The drupes with one stone. Fruit 1 seeded. Seeds endospermic. Endosperm oily. Cotyledons 2. Embryo chlorophyllous (1/1); straight to curved. Seedling. Germination phanerocotylar, or cryptocotylar. Physiology, biochemistry. Cyanogenic. Alkaloids present, or absent. Iridoids detected; ‘Route I’ type (normal and seco). Verbascosides detected (Cassinopsis). Proanthocyanidins present, or absent; when present, cyanidin. Flavonols present, or absent; quercetin. Ellagic acid absent (genera). Saponins/sapogenins present, or absent. Aluminium accumulation demonstrated. Geography, cytology. Sub-tropical to tropical. Pantropical, plus South Africa and Eastern Australia. X = 10, 11. Taxonomy. Subclass Dicotyledonae; at least mostly Tenuinucelli. Dahlgren’s Superorder Corniflorae; Cornales. Cronquist’s Subclass Rosidae; Celastrales. APG 3 core angiosperms; core eudicot; Superorder Asteranae; lamiid, or campanulid (Stemonuraceae only); Order Garryales, or Aquifoliales (Stemonuraceae only). Species 400. Genera 52; Alsodeiopsis, Apodytes, Calatola, Cantleya, Casimirella, Cassinopsis, Chlamydocarya, Citronella, Codiocarpus, Dendrobangia, Desmostachys, Discophora, Emmotum, Gastrolepis, Gomphandra, Gonocaryum, Grisollea, Hartleya, Hosiea, Icacina, Iodes, Irvingbaileya, Lasianthera, Lavigeria, Leretia, Mappia, Mappianthus, Medusanthera, Merrilliodendron, Miquelia, Natsiatopsis, Natsiatum, Nothapodytes, Oecopetalum, Ottoschulzia, Phytocrene, Pittosporopsis, Platea, Pleurisanthes, Polycephalium, Polyporandra, Poraqueiba, Pseudobotrys, Pyrenacantha, Rhaphiostylis, Rhyticaryum, Sarcostigma, Stachyanthus, Stemonurus, Whitmorea. This family exemplifies the well known difficulties in distributing certain Dicot families between Dahlgren’s Araliiflorae and Corniflorae. It had proved equally hard to assign them with confidence to the higher level groupings Crassinucelli and Tenuinucelli. This is interesting, given that the latter evidently represent a major divergence in the Dicot line of descent (cf.Young and Watson 1970, Chase et al. 1993). Economic uses, etc. A few cultivated ornamentals (Citronella (Villaresia). Illustrations. |
Support our developers





{kind=link}
{kind=link}