Family Lauraceae
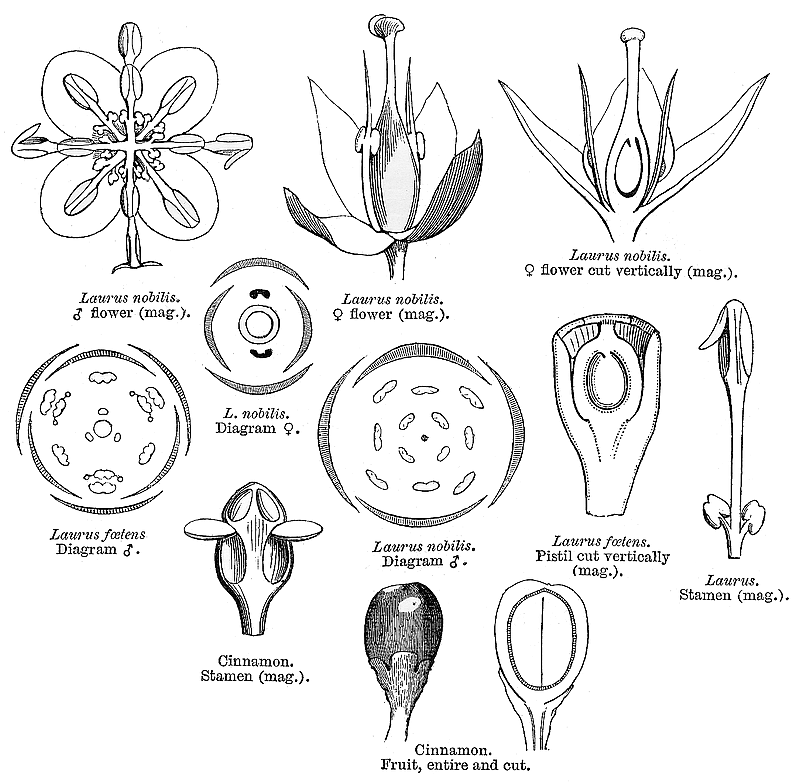
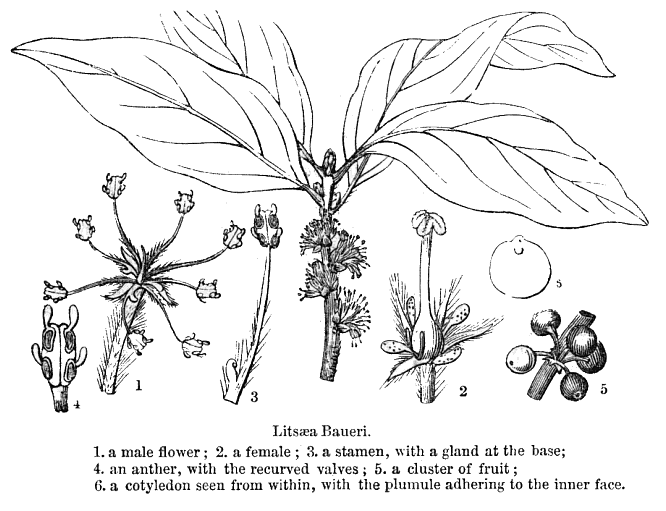
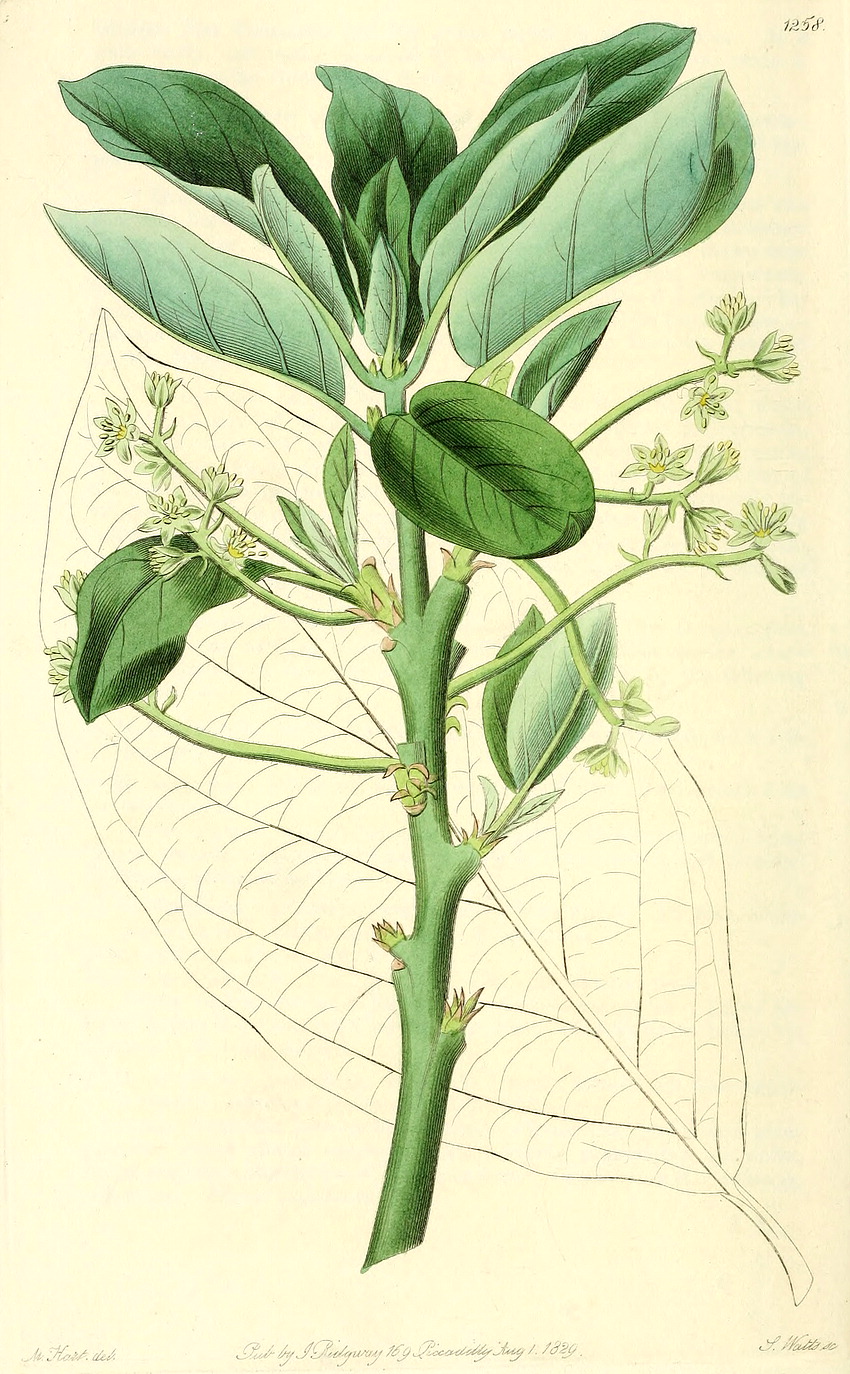
Lauraceae Juss.Including Perseaceae Horan.Excluding Cassythaceae Habit and leaf form. Trees and shrubs; bearing essential oils, or without essential oils. ‘Normal’ plants. Leaves well developed (usually). Plants autotrophic. The herbs perennial. Self supporting. Leptocaul. Mesophytic. Leaves nearly always evergreen; alternate (usually), or opposite (rarely), or whorled (rarely); usually spiral; leathery; petiolate; non-sheathing; gland-dotted; aromatic; simple. Lamina entire (usually), or dissected (lobed in e.g. Sassafras); when lobed, pinnatifid; pinnately veined; cross-venulate. Leaves exstipulate; without a persistent basal meristem. Domatia occurring in the family (14 genera); manifested as pits (mostly), or pockets, or hair tufts. Leaf anatomy. Stomata absent; mainly confined to one surface (abaxial); paracytic. Hairs present; mostly unicellular (and mostly thick walled). Adaxial hypodermis very often present. Lamina dorsiventral; without secretory cavities. The mesophyll usually with spherical etherial oil cells; containing mucilage cells, or not containing mucilage cells (less common than oil cells). Minor leaf veins without phloem transfer cells (Cinnamomum, Laurus, Persea). Stem anatomy. Cork cambium present; initially superficial (usually), or deep-seated. Nodes unilacunar (with two traces). Internal phloem absent. Secondary thickening developing from a conventional cambial ring. ‘Included’ phloem absent. Xylem with tracheids, or without tracheids; with fibre tracheids, or without fibre tracheids; with libriform fibres, or without libriform fibres. Vessel end-walls scalariform, or simple, or scalariform and simple. Vessels without vestured pits. Wood partially storied (VPI), or not storied; parenchyma paratracheal. Sieve-tube plastids P-type, or S-type; when P-type type I (b). Reproductive type, pollination. Plants hermaphrodite, or monoecious, or dioecious, or polygamomonoecious. Inflorescence, floral, fruit and seed morphology. Flowers solitary (rarely), or aggregated in ‘inflorescences’. The ultimate inflorescence unit cymose, or racemose. Inflorescences axillary; often umbelliform; with involucral bracts (often), or without involucral bracts. The involucres accrescent, or non-accrescent. Flowers small; often fragrant; regular; usually 3 merous; cyclic. Free hypanthium present (well developed, like a calyx tube, at least after fertilization). Hypogynous disk present, or absent. Perianth with distinct calyx and corolla, or sepaline, or of ‘tepals’ (the two similar, more or less sepaloid whorls variously interpreted as ‘tepals’ or ‘sepals’, hence the clumsy complexity of what follows); 6 (usually), or 4; free (but on a well developed hypanthium); (1–)2(–3) whorled; when more than one whorl, isomerous; sepaloid to petaloid; similar in the two whorls; green, or white, or cream, or yellow; fleshy, or non-fleshy; persistent, or deciduous; accrescent, or non-accrescent. Calyx if the P so interpreted, (4–)6; usually 2 whorled; polysepalous (on the hypanthium); regular; imbricate. Corolla if the inner P whorl so interpreted, 3; 1 whorled; polypetalous; imbricate; regular; green, or white, or yellow; fleshy. Petals sessile. Androecium (3–)9(–26). Androecial members free of the perianth (on the hypanthium); all equal, or markedly unequal; free of one another; (1–)3 whorled, or 4 whorled. Androecium exclusively of fertile stamens, or including staminodes (the innermost members often staminodal, and sometimes there are paired, nectariferous ?staminodal appendages on the filaments). Staminodes internal to the fertile stamens (in 1–2 whorls). Stamens (3–)9(–26); diplostemonous to polystemonous (usually), or reduced in number relative to the adjacent perianth to isomerous with the perianth; somewhat laminar to petaloid (by expansion of the filament and connective), or filantherous. Filaments appendiculate (the appendages paired, nectariferous), or not appendiculate. Anthers basifixed; non-versatile; dehiscing by longitudinal valves (opening from base to apex), or dehiscing via pores (in Hexapora); introrse (usually), or introrse and extrorse (with the latter state confined to the third whorl); bilocular, or four locular; bisporangiate, or tetrasporangiate, or bisporangiate and tetrasporangiate. Endothecium developing fibrous thickenings (or the cell walls become much thickened). Anther epidermis persistent. Microsporogenesis successive. The initial microspore tetrads tetrahedral (usually, predominantly), or isobilateral, or T-shaped, or linear. Anther wall initially with more than one middle layer (two, plus an endothecium); of the ‘basic’ type. Tapetum amoeboid (mostly), or glandular (in several genera). Pollen grains nonaperturate; 2-celled (in 4 genera). Gynoecium 1 carpelled (ostensibly), or 3 carpelled (theoretically). The pistil 1 celled. Gynoecium ostensibly monomerous; of one carpel (or at least, ostensibly so); superior (usually), or inferior (Hypodaphnis). Carpel stylate; apically stigmatic; 1 ovuled. Placentation apical. Stigmas dry type; papillate; Group II type. Ovules pendulous; apotropous; with dorsal raphe; non-arillate; anatropous; bitegmic; crassinucellate. Outer integument not contributing to the micropyle. Endothelium at least usually not differentiated (dubiously present in one Beilschmiedia pendula). Embryo-sac development Polygonum-type. Polar nuclei fusing prior to fertilization. Antipodal cells formed, or not formed (then the three nuclei degenerating early); when formed, 3; proliferating (rarely), or not proliferating. Synergids commonly exhibiting filiform apparatus. Hypostase present (in a few genera only), or absent. Endosperm formation nuclear. Endosperm haustoria present, or absent; micropylar (where observed, in Cryprocarya). Embryogeny onagrad, or asterad, or piperad (or with no clear pattern). Fruit fleshy (usually), or non-fleshy (very rarely). The fruiting carpel indehiscent; drupaceous, or baccate (usually). Fruit enclosed in the fleshy receptacle, or enclosed in the fleshy hypanthium, or without fleshy investment; 1 seeded. Seeds non-endospermic. Embryo well differentiated. Cotyledons massive, occasionally ruminate. Embryo achlorophyllous (5/6); straight. Seedling. Germination phanerocotylar, or cryptocotylar. Physiology, biochemistry. Not cyanogenic. Alkaloids present (commonly), or absent. Iridoids not detected. Proanthocyanidins present (usually), or absent; when present, cyanidin. Flavonols present; kaempferol and quercetin. Ellagic acid absent (8 species, 7 genera). Arbutin absent. Saponins/sapogenins absent. Aluminium accumulation demonstrated. Sugars transported as sucrose (e.g. Miscanteca (= Licaria)), or as oligosaccharides + sucrose (e.g. Sassafras), or as sugar alcohols + oligosaccharides + sucrose (e.g. Cinnamomum). Inulin recorded. Geography, cytology. Temperate to tropical. Pantropical and subtropical, extending into the temperate regions. X = 12. Taxonomy. Subclass Dicotyledonae; Crassinucelli. Dahlgren’s Superorder Magnoliiflorae; Laurales. Cronquist’s Subclass Magnoliidae; Laurales. APG 3 core angiosperms; Superorder Magnolianae; Order Laurales. Species 2000–2500. Genera 54; Actinodaphne, Adenodaphne, Aiouea, Alseodaphne, Anaueria, Aniba, Apollonias, Aspidostemon, Beilschmiedia, Brassiodendron, Caryodaphnopsis, Chlorocardium, Cinnadenia, Cinnamomum, Clinostemon, Cryptocarya, Dehaasia, Dicypellium, Dodecadenia, Endiandra, Endlicheria, Eusideroxylon, Gamanthera, Hexapora, Hypodaphnis, Iteadaphne, Kubitzkia, Laurus, Licaria, Lindera, Litsea, Mezilaurus, Nectandra, Neocinnamomum, Neolitsea, Nothaphoebe, Ocotea, Paraia, Persea, Phoebe, Phyllostemonodaphne, Pleurothyrium, Potameia, Potoxylon, Povedadaphne, Ravensara, Rhodostemonodaphne, Sassafras, Syndiclis, Triadodaphne, Umbellularia, Urbanodendron, Williamodendron. Economic uses, etc. Some important economic plants, including fruit from Persea americana (Avocado pear), cinnamon and camphor from Cinnamomum spp., aromatic oils oils from Lindera (benzoin) and Sassafras, and fragrant woods used in cabinet-making. Illustrations. |
Support our developers





{kind=link}
{kind=link}
{kind=link}