Family Loganiaceae
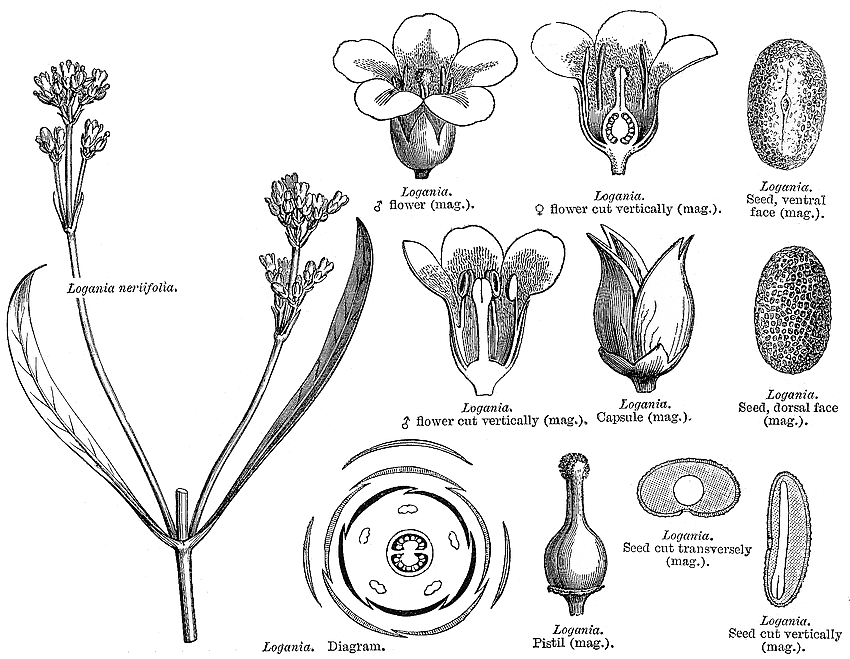
Loganiaceae Mart.~ Gentianaceae Excluding Antoniaceae, Buddlejaceae, Gelsemiaceae, Geniostomaceae, Mitreolaceae, Plocospermataceae, Potaliaceae, Spigeliaceae, Strychnaceae Habit and leaf form. Herbs, or shrubs (or subshrubs); non-laticiferous and without coloured juice. ‘Normal’ plants, or switch-plants (e.g. L. tortuosa); the switch forms with the principal photosynthesizing function transferred to stems. Leaves well developed, or much reduced. Annual (often, sometimes tiny), or perennial. Self supporting, or climbing. Mesophytic. Leaves opposite; ‘herbaceous’, or leathery, or membranous; petiolate to sessile; connate (commonly, more or less, via the stipular sheath, which is conspicuous even in the almost ‘leafless’ L. nuda), or not connate; simple. Lamina entire; linear, or lanceolate, or oblong, or ovate; one-veined, or pinnately veined; cross-venulate, or without cross-venules. Leaves more or less stipulate, or exstipulate (but then the opposing leaf bases connected by lateral lines). Stipules interpetiolar (usually represented by a ‘stipular sheath’, but each lateral component of the sheath is sometimes resolvable into a pair of joined stipules. Occasionally (e.g. L. serpyllifolia) the opposing stipules are completely concrescent to form conspicuous, free petiole-like structures which constitute a leaves-plus-stipules whorl at each node — cf. Rubiaceae); free of one another, or concrescent; with colleters, or without colleters; often much reduced. Leaf anatomy. Extra-floral nectaries absent. Mucilaginous epidermis present, or absent. Hairs present, or absent; eglandular. Complex hairs absent. Lamina dorsiventral, or isobilateral. The mesophyll with sclerencymatous idioblasts, or without sclerenchymatous idioblasts. Stem anatomy. Young stems cylindrical. Cork cambium present; initially deep-seated, or superficial. Nodes unilacunar (with 1–several traces), or tri-lacunar, or multilacunar. Primary vascular tissue bicollateral. Internal phloem present. Secondary thickening developing from a conventional cambial ring, or anomalous; from a single cambial ring. ‘Included’ phloem present, or absent. Xylem with tracheids, or without tracheids; with fibre tracheids, or without fibre tracheids; with libriform fibres, or without libriform fibres. Vessel end-walls simple (usually), or scalariform. Vessels without vestured pits. Wood ring porous (L. albiflora); not storied; parenchyma apotracheal, or paratracheal (very variable, sometimes very rare or absent). Reproductive type, pollination. Plants hermaphrodite, or dioecious. Female flowers with staminodes, or without staminodes. Gynoecium of male flowers pistillodial. Plants homostylous. Inflorescence, floral, fruit and seed morphology. Flowers aggregated in ‘inflorescences’ (usually), or solitary; in cymes, or in panicles (when not solitary). The ultimate inflorescence unit usually cymose. Inflorescences terminal, or axillary. Flowers bracteate; bracteolate; fragrant (sometimes), or odourless (?); regular; 4 merous (only L. micrantha), or 5 merous; cyclic; tetracyclic. Hypogynous disk small or absent. Perianth with distinct calyx and corolla; 8, or 10 (usually); 2 whorled; isomerous. Calyx 4 (rarely), or 5; 1 whorled; gamosepalous; blunt-lobed, or toothed; regular; imbricate; when K 5, with the median member anterior. Corolla 4 (rarely), or 5; 1 whorled; gamopetalous (often internally hairy); imbricate; (tubular-) campanulate, or hypocrateriform, or rotate; regular; white, or yellow, or pink. Androecium 4 (rarely), or 5. Androecial members adnate (epipetalous); free of one another; 1 whorled. Androecium of male-fertile flowers, exclusively of fertile stamens. Stamens 4 (L. micrantha), or 5; inserted midway down the corolla tube, or in the throat of the corolla tube; isomerous with the perianth; oppositisepalous; alternating with the corolla members. Filaments not appendiculate. Anthers dorsifixed; dehiscing via longitudinal slits (glabrous); introrse; unappendaged. Pollen grains aperturate; 3 aperturate; colporate. Gynoecium 2 carpelled. Carpels reduced in number relative to the perianth. The pistil 2 celled. Gynoecium syncarpous; eu-syncarpous; superior. Ovary 2 locular. Gynoecium median. Ovary sessile. Gynoecium stylate. Styles 1; attenuate from the ovary; apical. Stigmas 1; 1 lobed, or 2 lobed; clavate, or capitate. Placentation axile (the placentas not bilobed). Ovules differentiated; (1–)3–50 per locule ((one to) several to ‘many’); anatropous, or hemianatropous; unitegmic. Fruit non-fleshy; dehiscent, or a schizocarp (the carpels becoming more or less free, dehiscing longitudinally to the inside along the septum). Mericarps when deemed schizocarpic, 2; comprising follicles. Fruit if not schizocarpic, a capsule. Capsules denticidal, septicidal, and loculicidal (dehiscing only in the upper half, becoming mitre-shaped). Fruit 1–100 seeded (‘several to many’). Seeds endospermic; wingless (and not flattened). Cotyledons 2. Embryo straight. Seedling. Germination phanerocotylar. Physiology, biochemistry. Not cyanogenic. Alkaloids present (mostly), or absent. Iridoids detected; ‘Route I’ type (normal and seco). Verbascosides not detected. Cornoside not detected. Proanthocyanidins absent. Ellagic acid absent. Saponins/sapogenins present, or absent. Aluminium accumulation not found. Peculiar feature. Lamina tip not abaxially pouched. Geography, cytology. Paleotropical, Australian, and Antarctic. Temperate (a few), or sub-tropical to tropical. Australia, New Caledonia and New Zealand. 2n = 32. Supposed basic chromosome number of family: 8. Ploidy levels recorded: 4. Taxonomy. Subclass Dicotyledonae; Tenuinucelli. Dahlgren’s Superorder Gentianiflorae; Gentianales. Cronquist’s Subclass Asteridae; Gentianales. APG 3 core angiosperms; core eudicot; Superorder Asteranae; lamiid; Order Gentianales. Species about 30. Genera 1, or 3; Logania. Leeuwenberg (1980) provided detailed descriptive data for the genera of Loganiaceae sensu lato. Struwe et al. (1994) applied cladistics to a sample comprising that assemblage plus small samples from assorted other tenuinucellate families, and published a classification described by Leeuwenberg (1997) as ‘fraught with pretentious superficiality’. In addition to ignoring intra-taxon variation, as exemplified by him, their sparse data (presented encoded in almost unreadable and accident prone form) represent only a very perfunctory contribution towards preparing functional descriptions of new and recircumscribed families. In no way do they approach the standards which are required for this package, and which are essential for convincing assessments of relationships. The detailed descriptions attempted here require further pursuit of available literature, notably re embryological and anther developmental characters, and various morphological characters of apparent classificatory importance (e.g. flower and gynoecium orientation, directions of anther dehiscence, gynoecium and fruit details), for all the seggregates of the traditional Loganiaceae (cf. Antoniaceae, Buddlejaceae, Gelsemiaceae, Geniostomaceae, Mitreolaceae, Potaliaceae, Spigeliaceae), Strychnaceae). Meanwhile, in terms of these extended descriptions, Logania is indeed incompletely separable from Gentianaceae (cf. Struwe et al.), but differs from their Geniostomaceae only in its colporate pollen and wood vessels without vestured pits. It seems to differ from Mitreolaceae (q.v., the other members of Struwe and Albert’s Loganicaceae) in the presence of bracteoles, in the consistently imbricate corolla, and in the consistent absence of synstyly. Illustrations. |
Support our developers





{kind=link}
{kind=link}