Family Meliaceae
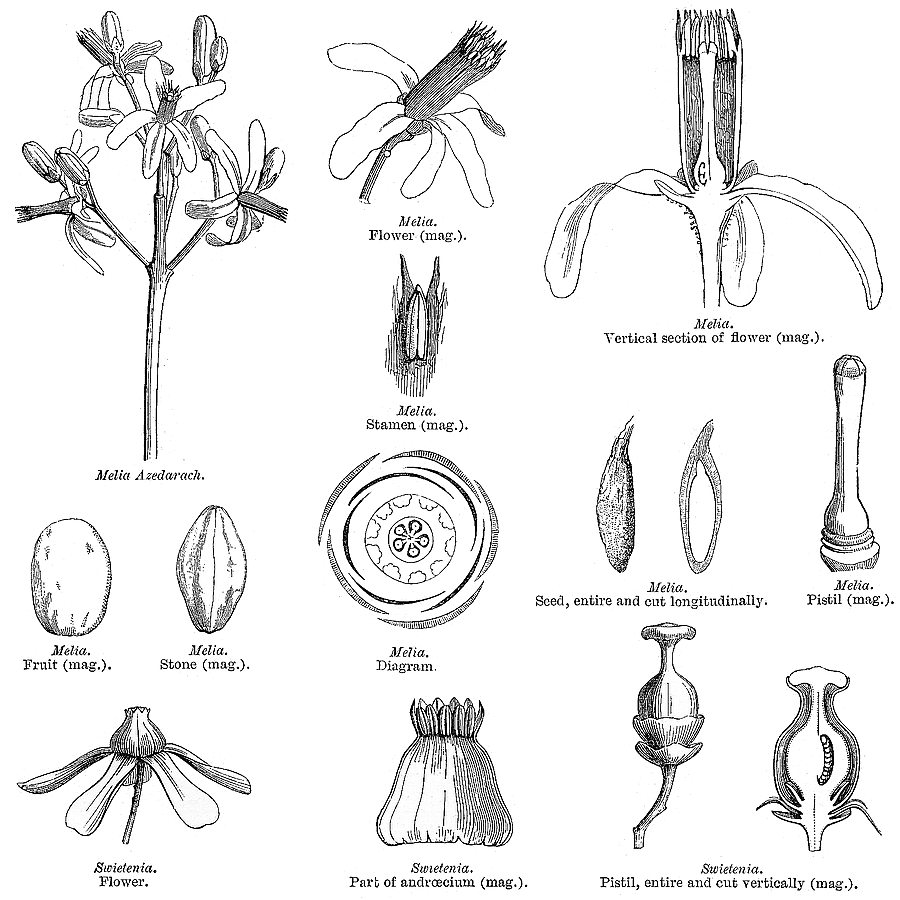
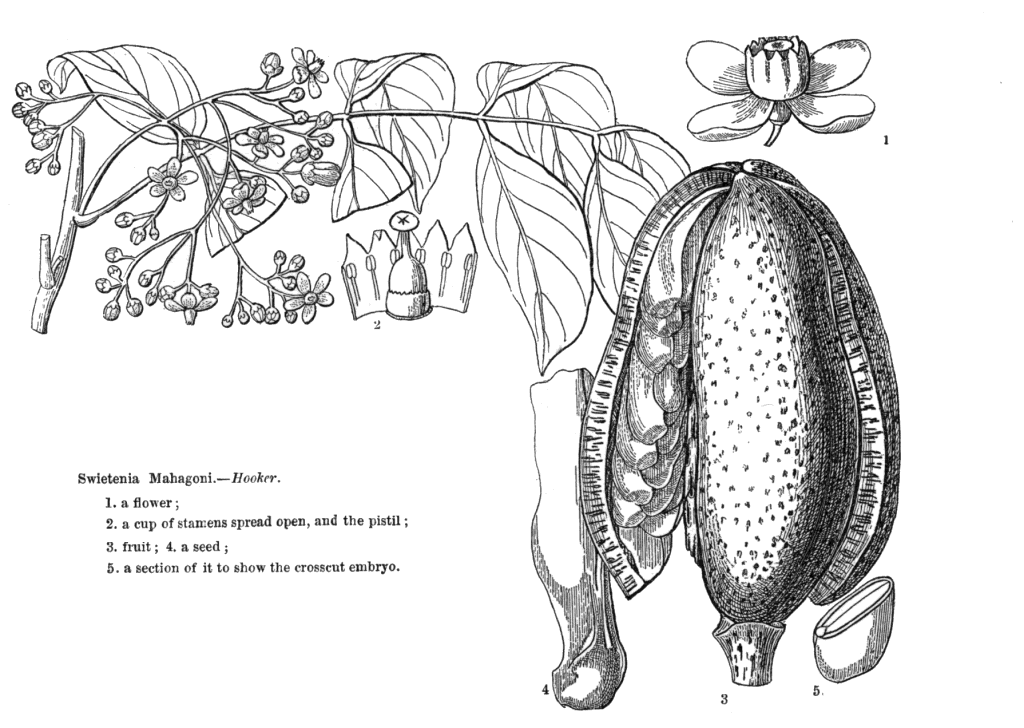
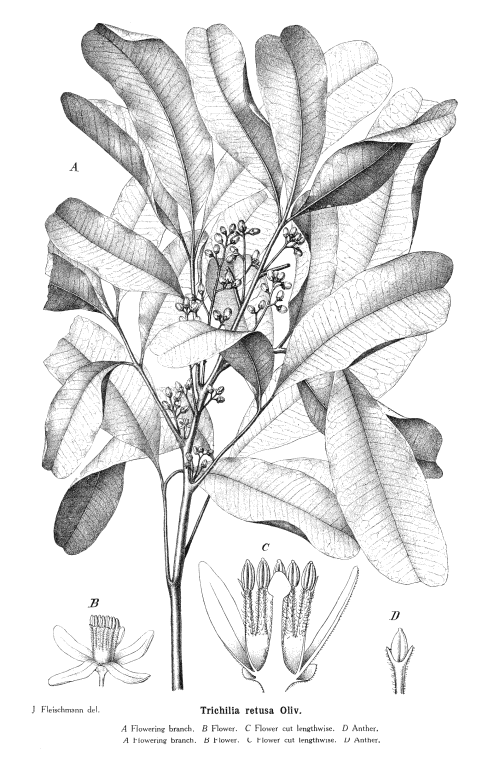
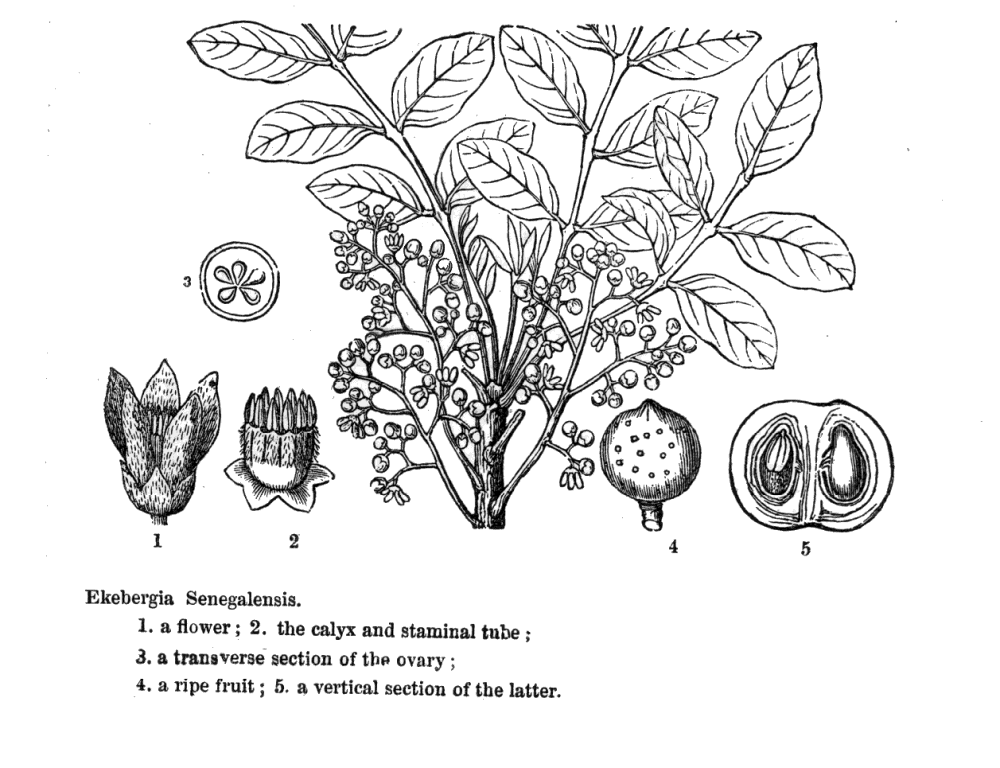
Meliaceae Juss.Including Cedrelaceae R. Br.Excluding Aitoniaceae, Flindersiaceae Habit and leaf form. Trees, or shrubs (or suckering shrublets), or herbs (rarely, e.g. Naregamia); laticiferous (rarely, with milky juice exuding from the bark), or with coloured juice; bearing essential oils. Stem growth conspicuously sympodial (rarely), or not conspicuously sympodial. Often pachycaul. Mesophytic. Leaves alternate (nearly always), or opposite (rarely, decussate); nearly always spiral; petiolate; non-sheathing; not gland-dotted; compound (usually), or simple; pinnate (mostly, pari- or imparipinnate), or unifoliolate (rarely), or ternate (rarely), or bifoliolate (very rarely), or bipinnate (very rarely). Lamina pinnately veined; cross-venulate. Leaves exstipulate. Lamina margins entire (usually), or crenate, or serrate, or dentate (or lobed). Vegetative buds scaly, or not scaly. Leaves without a persistent basal meristem. Domatia occurring in the family (6 genera); manifested as pits, or pockets, or hair tufts. Leaf anatomy. Mucilaginous epidermis present, or absent. The mesophyll with sclerencymatous idioblasts, or without sclerenchymatous idioblasts. Minor leaf veins without phloem transfer cells (Turraea). Stem anatomy. Cork cambium present; initially superficial. Nodes mostly penta-lacunar. Internal phloem absent. Secondary thickening developing from a conventional cambial ring. The secondary phloem stratified into hard (fibrous) and soft (parenchymatous) zones, or not stratified. ‘Included’ phloem absent. Xylem with fibre tracheids, or without fibre tracheids; with libriform fibres, or without libriform fibres (the fibres septate or non-septate); with vessels. Vessel end-walls simple. Vessels without vestured pits. Wood diffuse porous (usually), or ring porous to semi-ring porous; storied; parenchyma apotracheal, or paratracheal. Reproductive type, pollination. Plants hermaphrodite (usually), or monoecious, or dioecious, or polygamomonoecious. Gynoecium of male flowers pistillodial, or vestigial (present as well developed rudiments, perhaps important in pollination). Pollination usually) entomophilous; via hymenoptera, or via lepidoptera; mechanism conspicuously specialized (passive pollen presenters, in at least three genera), or unspecialized. Inflorescence, floral, fruit and seed morphology. Flowers solitary, or aggregated in ‘inflorescences’; in cymes, in racemes, in panicles, and in spikes. The ultimate inflorescence unit cymose (usually, thyrsoid), or racemose. Inflorescences axillary (usually), or terminal, or leaf-opposed, or epiphyllous, or cauliflorous; usually paniculate with cymose branchlets (thyrsoid), less often racemose, fasciculate or spicate, or flowers paired or solitary. Flowers minute to large; calyptrate, or not calyptrate; regular; cyclic; tetracyclic to polycyclic. Floral receptacle with neither androphore nor gynophore. Free hypanthium absent. Hypogynous disk present, or absent; when present, intrastaminal; variable in form, stipitate, patelliform, cyathiform or tubular, free from or more or less fused with the bases of the staminal tube or ovary. Perianth with distinct calyx and corolla; (5–)6–12(–21); 2 whorled, or 3 whorled (the corolla sometimes in two whorls); isomerous, or anisomerous. Calyx (2–)3–5(–7) (sometimes transitional to bracteoles); 1 whorled; gamosepalous (usually), or polysepalous; entire (occasionally, truncate or closed and circumcissile), or lobulate to blunt-lobed; regular; calyptrate (rarely), or not calyptrate; imbricate, or open in bud, or valvate (rarely, or almost closed when circumcissile). Corolla 3–7(–14); 1 whorled, or 2 whorled; not appendiculate; polypetalous, or gamopetalous (rarely basally united, but often fused with the staminal tube); imbricate, or contorted, or valvate (less often); not fleshy. Androecium (3–)5–10(–23), or 10–100 (to ‘many’, if appendages on the tube are treated as staminodial). Androecial members unbranched; free of the perianth, or adnate (with the staminal tube adnate to the corolla); free of one another, or coherent (usually forming a staminal tube, which may be complete or incomplete, short or up to 14 cm long, globose, urceolate, campanulate, etc., or cylindrical and then sometimes curved or inflated distally, and which free or partially united appendages); when joined, 1 adelphous; 1–21 whorled (usually haplostemonous, sometimes with two stamen whorls, and up to many series of ‘appendages’). Androecium exclusively of fertile stamens, or including staminodes (the staminal tube being entire, crenate, lobed or bearing appendages). Staminodes when present, 5–100 (as many as the stamens, or twice the number, rarely up to ten times the number). Stamens (3–)5–10(–23); isomerous with the perianth, or diplostemonous; usually alternating with appendages, sometimes opposite them; filantherous, or petaloid, or petaloid and with sessile anthers (often with the anthers sessile on the corolla-like staminal tube). Anthers more or less dorsifixed; versatile; dehiscing via longitudinal slits; introrse; tetrasporangiate; unappendaged. Endothecium developing fibrous thickenings. Anther epidermis persistent. Microsporogenesis simultaneous. The initial microspore tetrads tetrahedral, or isobilateral, or decussate. Anther wall initially with more than one middle layer (2 or 3). Tapetum glandular. Pollen shed in aggregates (rarely), or shed as single grains; rarely in tetrads. Pollen grains aperturate; 2–5 aperturate; colporate (sometimes slightly ruporate); 2-celled (in 4 genera), or 3-celled (in Azadirachta only). Gynoecium (1–)2–6(–20) carpelled. Carpels reduced in number relative to the perianth to increased in number relative to the perianth. The pistil (1–)2–6(–20) celled. Gynoecium syncarpous; synstylovarious, or eu-syncarpous; superior. Ovary (1–)2–6(–20) locular (reflecting the carpel number). Gynoecium non-stylate to stylate. Styles when demarcated, 1; when perceptible, attenuate from the ovary, or from a depression at the top of the ovary; apical. Stigmas 1; variously clavate, or capitate, or subpeltate to peltate (commonly forming a conspicuous receptaculum pollinis); wet type; papillate; Group III type. Placentation when unilocular (i.e. occasionally), parietal; usually axile. Ovules in the single cavity when unilocular 1, or 2, or 3–100 (i.e. one to ‘many’); 1–2 per locule (Melioideae), or 3–50 per locule (i.e. one to ‘many’, in Swietenioideae); usually pendulous; epitropous (micropyle superior); with ventral raphe; collateral, or superposed, or biseriate; arillate (Melioideae), or non-arillate (Swietenioideae); orthotropous, or anatropous, or campylotropous; bitegmic; crassinucellate. Outer integument contributing to the micropyle, or not contributing to the micropyle. Embryo-sac development Polygonum-type. Polar nuclei fusing prior to fertilization. Antipodal cells formed; 3; not proliferating; ephemeral. Synergids hooked (sometimes with filiform apparatus). Endosperm formation nuclear. Embryogeny onagrad. Fruit fleshy, or non-fleshy; dehiscent, or indehiscent; a capsule, or a berry, or a drupe, or a nut (rarely); without fleshy investment. Capsules septicidal, or loculicidal. Seeds endospermic (rarely), or non-endospermic; with a testa; winged (Swietenioideae), or wingless. Embryo well differentiated. Cotyledons 2. Embryo chlorophyllous (5/6), or achlorophyllous (3/4); straight, or curved. Seedling. Germination phanerocotylar, or cryptocotylar. Physiology, biochemistry. Cyanogenic (?), or not cyanogenic. Alkaloids present (commonly), or absent. Iridoids detected (doubtfully, in Xylocarpus), or not detected (see Jensen 1991). Proanthocyanidins present, or absent; cyanidin. Flavonols present; quercetin, or kaempferol and quercetin. Ellagic acid absent (4 species, 3 genera). Saponins/sapogenins present (rarely), or absent. Aluminium accumulation not found. Sugars transported as sucrose (e.g. in Cedrela), or as oligosaccharides + sucrose (in four other genera). Geography, cytology. Tropical (mostly), or sub-tropical (few). Pantropical to subtropical and warm. X = 10–14(+). Taxonomy. Subclass Dicotyledonae; Crassinucelli. Dahlgren’s Superorder Rutiflorae; Rutales. Cronquist’s Subclass Rosidae; Sapindales. APG 3 core angiosperms; core eudicot; Superorder Rosanae; malvid; Order Sapindales. Species 575. Genera about 50; Aglaia, Anthocarapa, Aphanamixis, Astrotrichilia, Azadirachta, Cabralea, Calodecarya, Capuronianthus, Carapa, Cedrela, Chisocheton, Chukrasia, Cipadessa, Dysoxylum, Ekebergia, Entandrophragma, Guarea, Heckeldora, Humbertioturraea, Khaya, Lansium, Lepidotrichilia, Lovoa, Malleastrum, Melia, Munronia, Naregamia, Neobeguea, Owenia, Pseudobersama, Pseudocarapa, Pseudocedrela, Pterorhachis, Reinwardtiodendron, Ruagea, Sandoricum, Schmardaea, Soymida, Sphaerosacme, Swietenia, Synoum, Toona, Trichilia, Turraea, Turraeanthus, Vavaea, Walsura, Xylocarpus. Pennington and Styles 1975. Economic uses, etc. Edible fruit from Lansium domesticum (langsat, ayer-ayer); timber from Swietenia (mahogany), Kaha (African mahogany). Illustrations. |
Support our developers





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}