Family Chenopodiaceae
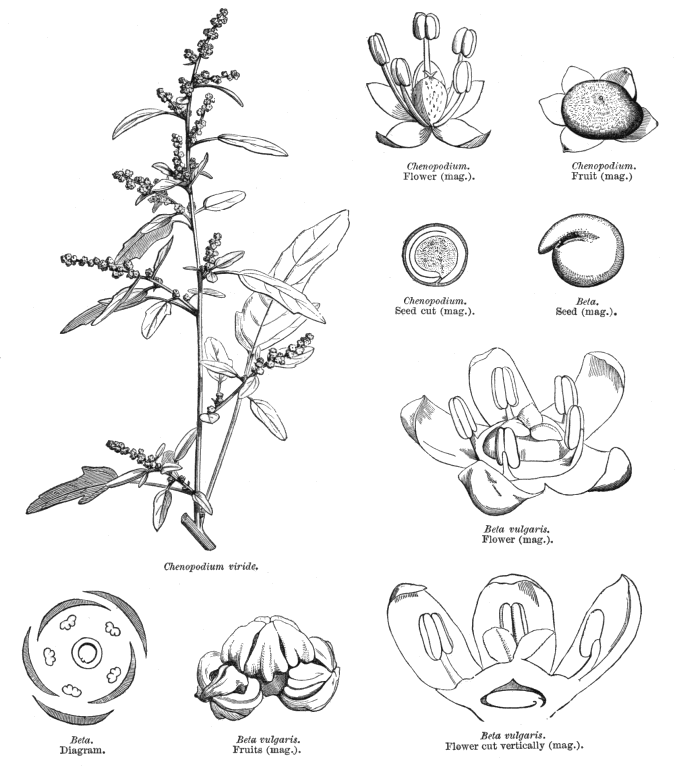
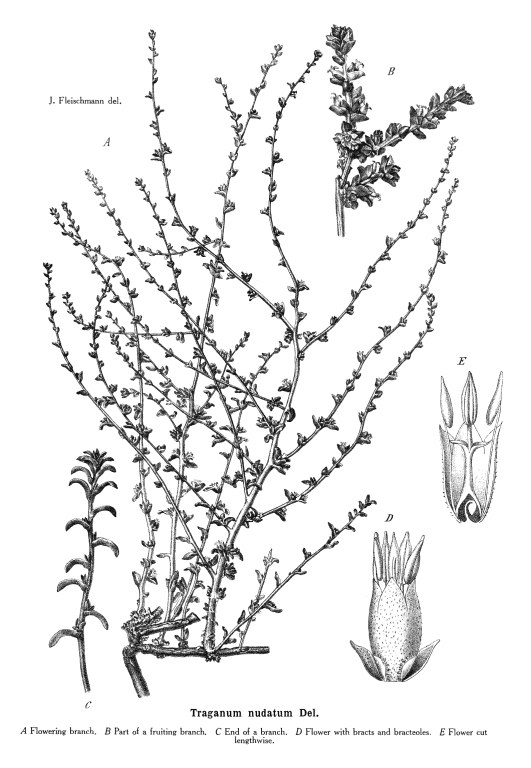
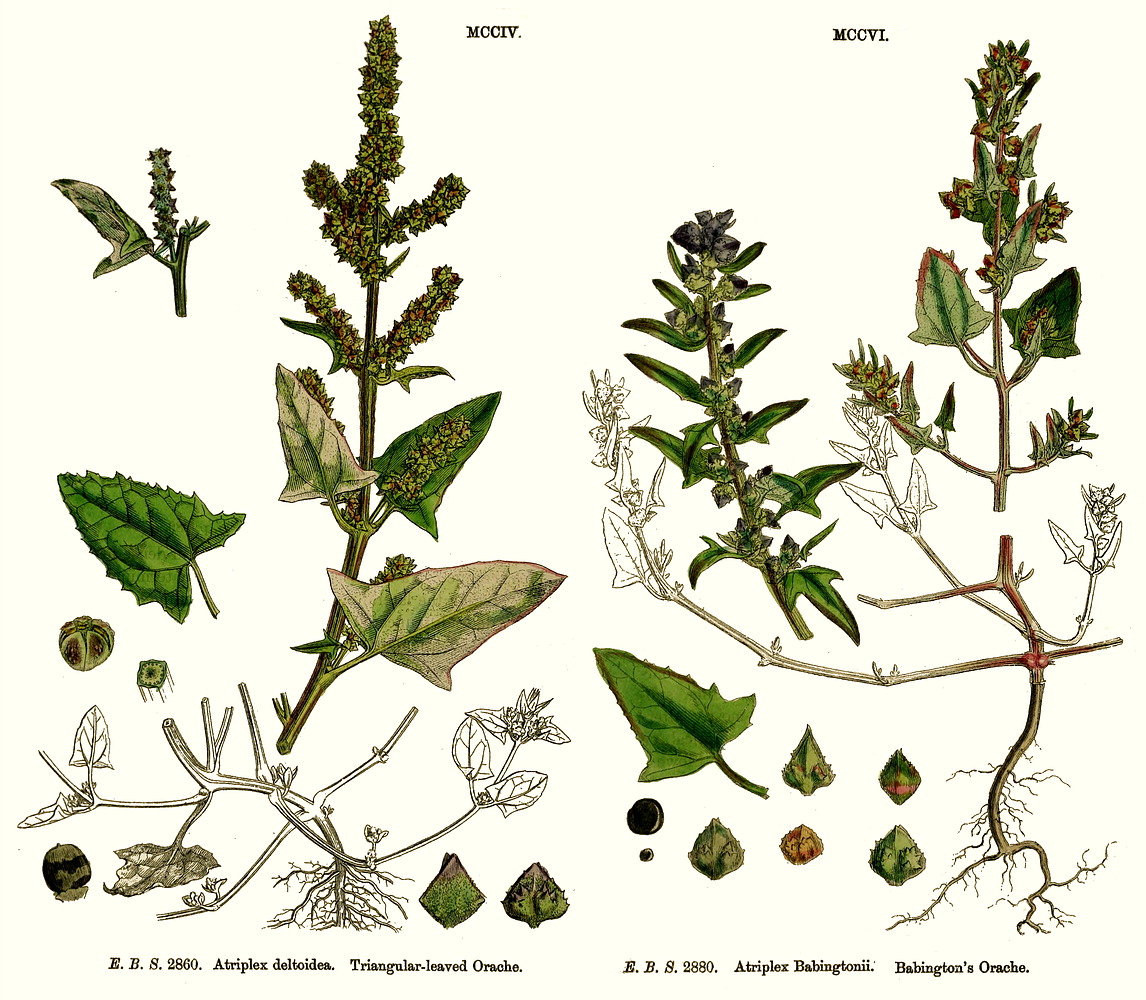
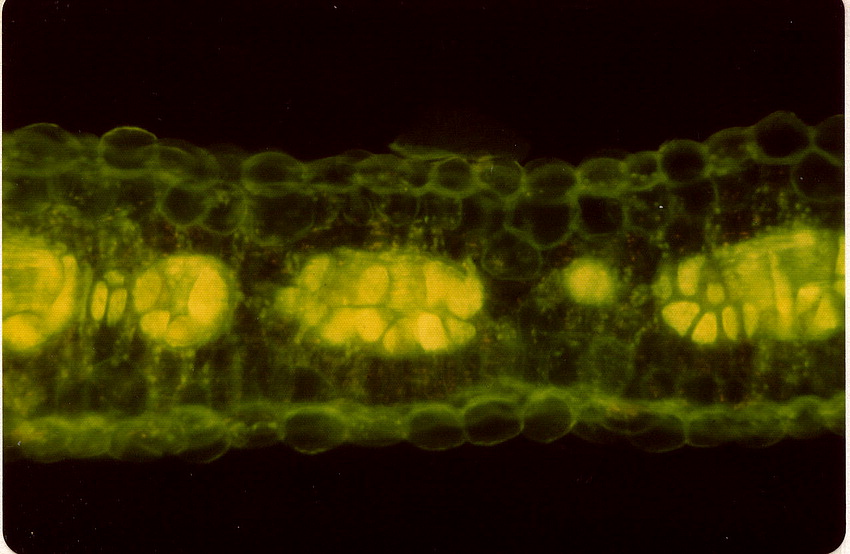
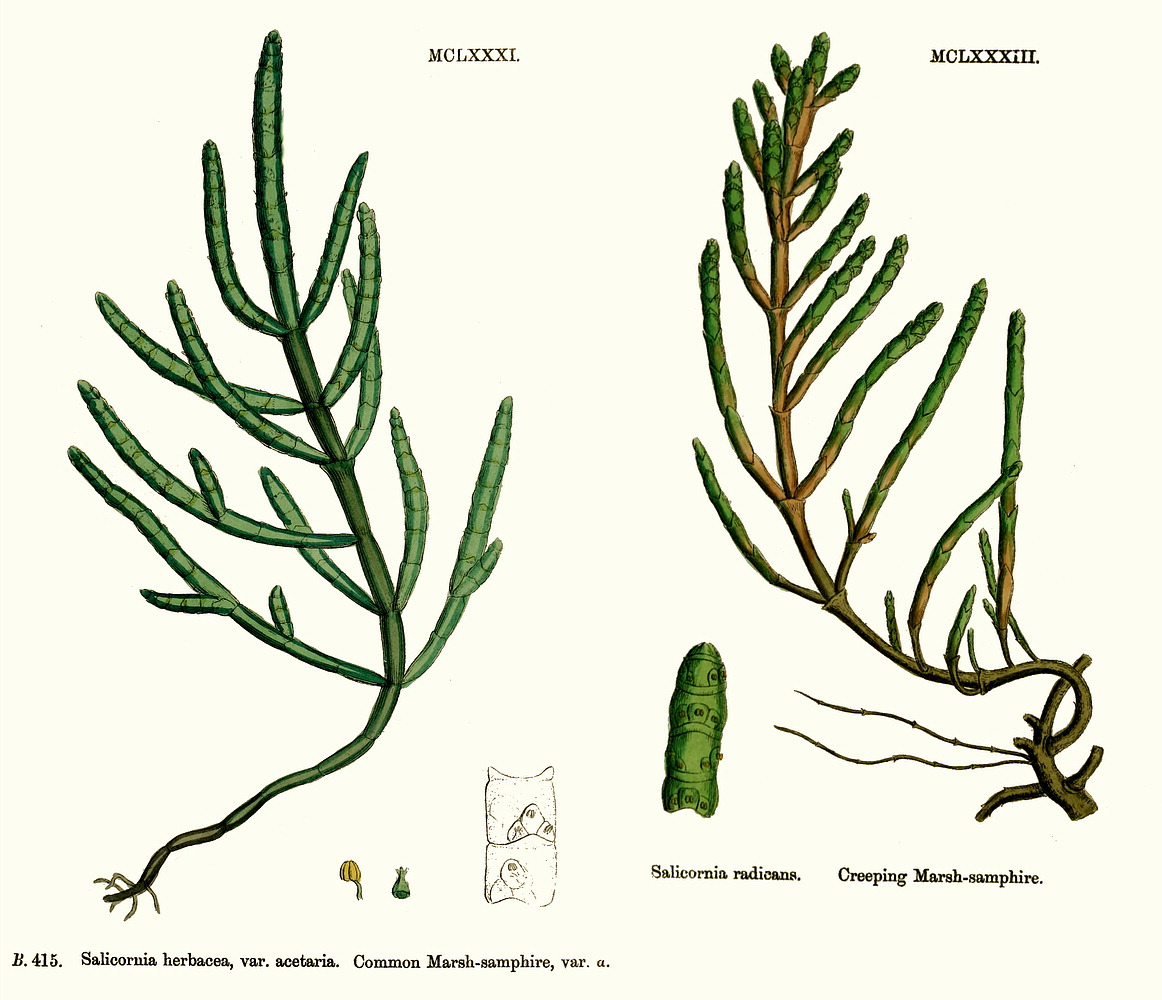
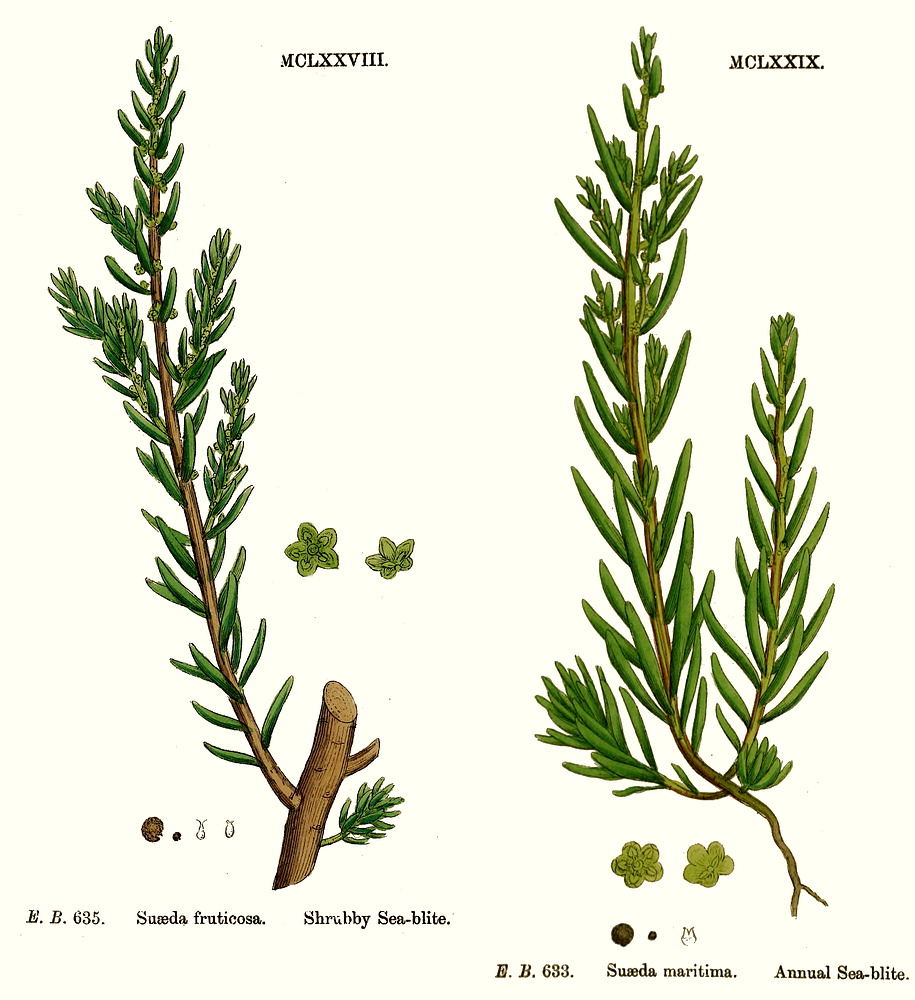
Chenopodiaceae Vent.~ Amaranthaceae, according to APG (1998) Including Atriplicaceae Juss., Corispermaceae Link, Farinaceae Dulac, Salicornieae (Salicorniaceae) J.G. Agardh, Salsolaceae Moq.-TandonExcluding Dysphaniaceae, Halophytaceae, Sarcobataceae Habit and leaf form. Herbs (mostly), or shrubs (some), or trees (few, small), or lianas (few). ‘Normal’ plants and switch-plants; sometimes more or less ‘cactoid’ (e.g. Salicornia). Leaves well developed, or much reduced, or absent. Plants with roots; succulent, or non-succulent. Annual, or biennial, or perennial (often glaucous); with a basal aggregation of leaves, or with neither basal nor terminal aggregations of leaves. Self supporting (usually), or climbing (few). Helophytic to xerophytic (nearly all halophytic). Leaves minute to large; alternate, or opposite; when alternate, spiral, or distichous (rarely); ‘herbaceous’, or fleshy, or membranous; petiolate to sessile; when opposite, connate (not uncommonly, e.g. represented by lobed cupules at the internodes of succulent stems), or not connate; sheathing, or non-sheathing; simple; epulvinate. Lamina dissected, or entire; one-veined (?), or pinnately veined; hastate, or sagittate, or attenuate at the base, or cuneate at the base. Leaves exstipulate; without a persistent basal meristem. General anatomy. Plants with ‘crystal sand’, or without ‘crystal sand’. Leaf anatomy. Hydathodes present (sometimes), or absent. Minor leaf veins without phloem transfer cells (6 genera). Stem anatomy. Cork cambium present; initially deep-seated, or superficial. Nodes unilacunar. Cortical bundles absent. Medullary bundles present (commonly), or absent. Internal phloem absent. Secondary thickening developing from a conventional cambial ring, or anomalous; when anomalous, via concentric cambia (very commonly), or from a single cambial ring (?). ‘Included’ phloem present (very commonly), or absent. Xylem with libriform fibres; with vessels. Vessel end-walls simple. Wood not storied (VP, in all those with secondary growth); parenchyma paratracheal (and conjunctive). Sieve-tube plastids P-type; type III (c). Reproductive type, pollination. Plants hermaphrodite (usually), or monoecious (sometimes), or andromonoecious (rarely), or gynomonoecious, or dioecious (rarely), or polygamomonoecious. Inflorescence, floral, fruit and seed morphology. Flowers solitary (then axillary), or aggregated in ‘inflorescences’ (or sometimes paired); when solitary or paired, axillary; when flowers aggregated, in cymes (the cymes mostly in racemose arrangements), or in catkins (male inflorescences of Sarcobatus). The ultimate inflorescence unit cymose. Inflorescences terminal, or axillary; mostly racemes of cymes. Flowers minute, or small; regular; cyclic; when hermaphrodite, usually tricyclic (sometimes 2-cyclic). Free hypanthium present (the stamens then inserted ‘on a disk on the calyx’), or absent. Hypogynous disk often present (but not always?). Perianth sepaline, or vestigial to absent; (1–)5(–6); joined (imbricate); 1 whorled; fleshy, or non-fleshy; persistent; accrescent, or non-accrescent. Calyx (interpreting the perianth as such) (1–)3–5(–6); gamosepalous (nearly always), or polysepalous (in Salsola?); commonly becoming appendaged with spines, tubercles or wings; fleshy (commonly), or non-fleshy (then usually herbaceous or membranous); persistent (in the fruit); accrescent (usually, enclosing the fruit and often appendaged with wings, tubercles or spines), or non-accrescent; imbricate. Androecium 3–5. Androecial members free of the perianth, or adnate (to the base of the perianth); all equal (by contrast with most Amaranthaceae); free of one another (usually, by contrast with most Amaranthaceae), or coherent; when coherent 1 adelphous (connate at the base); 1 whorled. Androecium nearly always exclusively of fertile stamens (by contrast with many Amaranthaceae). Stamens 3–5; usually isomerous with the perianth; oppositisepalous (when ascertainable). Anthers bent inwards in bud; dehiscing via longitudinal slits; tetrasporangiate. Endothecium developing fibrous thickenings. Microsporogenesis simultaneous. The initial microspore tetrads tetrahedral, or isobilateral, or decussate. Anther wall initially with one middle layer, or initially with more than one middle layer; of the ‘monocot’ type. Tapetum glandular (usually), or amoeboid. Pollen grains aperturate; 8–90 aperturate; (oligo-to poly-) foraminate; spinulose; 3-celled (7 genera). Gynoecium (2–)5 carpelled. The pistil 1 celled. Gynoecium syncarpous; synovarious to synstylovarious; superior (usually), or partly inferior (rarely). Ovary 1 locular. Styles (1–)2–3(–4); usually partially joined. Stigmas dry type; papillate; Group II type. Placentation basal. Ovules in the single cavity 1; pendulous, or ascending; non-arillate; campylotropous; bitegmic; crassinucellate. Outer integument not contributing to the micropyle. Embryo-sac development Polygonum-type. Polar nuclei ‘usually’ fusing prior to fertilization. Antipodal cells formed; 3; proliferating (a little, e.g. Beta vulgaris), or not proliferating (usually); usually ephemeral. Synergids pear-shaped, or hooked. Endosperm formation nuclear. Endosperm haustoria present, or absent; when developed, chalazal. Embryogeny chenopodiad. Fruit non-fleshy; not an aggregate; indehiscent, or dehiscent; a nut (usually), or capsular-indehiscent (commonly a utricle); enclosed in the fleshy hypanthium, or enclosed in the fleshy perianth (commonly), or without fleshy investment. Gynoecia of adjoining flowers combining to form a multiple fruit, or not forming a multiple fruit. Seeds more or less non-endospermic. Perisperm present, or absent (the variation of classificatory interest). Seeds with starch. Cotyledons 2. Embryo chlorophyllous (5/11), or achlorophyllous (3/5 (Atriplex, Chenopodium, Beta)); coiled (usually), or curved, or bent. Seedling. Germination phanerocotylar. Physiology, biochemistry. Cyanogenic, or not cyanogenic. Alkaloids present, or absent. Iridoids not detected. Arthroquinones detected (Salsola); polyacetate derived. Proanthocyanidins absent. Flavonols present; kaempferol, or kaempferol and quercetin. Ellagic acid absent (7 species, 7 genera). Arbutin absent. Betalains present. Saponins/sapogenins present, or absent. Aluminium accumulation not found. Plants often accumulating free oxalates. Inulin recorded (? — Beta). C3, or C4. C3 physiology recorded directly in Agriophyllum, Allenrolfia, Anthochlamys, Arthrocnemum, Atriplex, Bassia, Beta, Blitum, Ceratocarpus, Chenopodium, Eurotia, Halimione, Halisarcia, Halocnemum, Halosarcia, Haplopeplis, Halostachya, Kalidium, Micronemum, Pachycornia, Salicornia, Salsola, Sclerostegia, Spinacia, Suaeda, Tecticornia. C4 physiology recorded directly in Aellinia, Agathophora, Anabasis, Arthrophytum, Atriplex, Bassia, Camphorosma, Climacoptera, Cornulaca, Gamanthus, Girgensohnia, Halanthium, Halarchon, Halimocnemis, Halocharis, Halogeton, Halosarcia, Halotis, Haloxylon, Hammada, Horaninovia, Hypocyclix, Kochia, Noaea, Panderia, Petrosimonia, Salsola, Suaeda, Theleophyton, Traganum. Anatomy C4 type (Aellenia, Anabasis, Arthrophytum, Atriplex, Bassia, Camphorosma, Chenolea, Girgensohnia, Halanthium, Halosarcia, Kochia, Panderia, Petrosimonia, Salsola, Suaeda, etc.), or non-C4 type (Allenrolfea, Arthrocnemum, Atriplex, Bassia, Ceratocarpus, Chenolea, Chenopodium, Halimione, Halocnemum, Halopeplis, Halosarcia, Kochia, Maireana, Pachycornia, Salicornia, Sarcocornia, Sclerostegia, Suaeda, Tecticornia etc.). Geography, cytology. Temperate to sub-tropical. Widespread, but missing from Central America, Asia, Malaysia and most of Africa. X = (6–)9. Taxonomy. Subclass Dicotyledonae; Crassinucelli. Dahlgren’s Superorder Caryophylliflorae; Caryophyllales. Cronquist’s Subclass Caryophyllidae; Caryophyllales. APG 3 core angiosperms; core eudicot; Superorder Caryophyllanae; Order Caryophyllales (under Amaranthaceae!). Species 1400. Genera about 105; Acroglochin, Aellenia (= Halothamnus), Agathophora, Agriophyllum, Alexandra, Allenrolfea, Anabasis, Anthochlamys, Aphanisma, Archiatriplex, Arthocnemum, Arthrophytum, Atriplex, Axyris, Baolia, Bassia, Beta, Bienertia, Blitum, Borsczowia, Camphorosma, Ceratocarpus, Chenopodium, Climacoptera, Corispermum, Cornulaca, Cremnophyton, Cyathobasis, Cycloloma, Didymanthus, Dissocarpus, Einadia, Enchylaena, Eremophea, Eriochiton, Exomis, Fadenia, Fredolia, Gamanthus, Girgensohnia, Grayia, Hablitzea, Halanthium, Halarchon, Halimione, Halimocnemis, Halocharis, Halocnemum, Halogeton, Halopeplis, Halosarcia, Halostachys, Halothamnus, Haloxylon, Hammada, Hemichroa (~ Amaranthaceae), Heterostachys, Holmbergia, Horaninovia, Iljinia, Kalidium, Kirilowia, Kochia, Krascheninnikovia, Lagenantha, Maireana, Malacocera, Manochlamys, Microcnemum, Microgynoecium, Monolepis, Nanophyton, Neobassia, Nitrophila, Noaea, Nucularia, Ofaiston, Oreobliton, Osteocarpum, Pachycornia, Panderia, Petrosimonia, Physandra, Piptoptera, Polycnemum, Rhagodia, Raphidophyton, Roycea, Salicornia, Salsola, Sarcocornia, Scleroblitum, Sclerochlamys, Sclerolaena, Sclerostegia, Seidlitzia, Sevada, Spinacia, Stelligera, Suaeda, Suckleya, Sympegma, Tecticornia, Tegicornia, Teloxys, Threlkeldia, Traganopsis, Traganum, Zuckia. Sarcobatus has long been acknowledged an an anomalous member of this family, e.g. by Bentham and Hooker (1880), who presented it as a monogeneric tribe. Behnke (1997) proposes raising it to family rank, because sieve-element plastid form supports recent chloroplast DNA sequencing studies in portraying it nearer Phytolaccaceae than Chenopodiaceae. He provides no organized comparative descriptive data, merely a short Latin ‘diagnosis’ of the new family, on the basis of which Sarcobatus is incompletely separable morphologically from the description of Chenopodiaceae given here and elsewhere, with no indication of how the latter should be modified. Economic uses, etc. Contributes sugarbeet and Swiss chard (Beta vulgaris), spinach (Spinacia oleracea), and ‘saltbush’ pastures. Illustrations. |
Support our developers





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}