Family Lecythidaceae
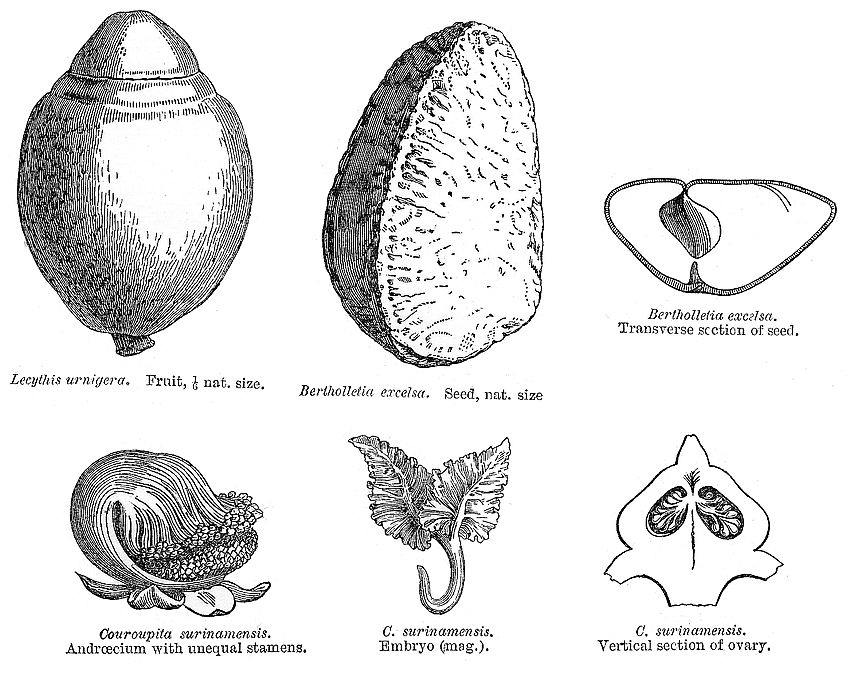
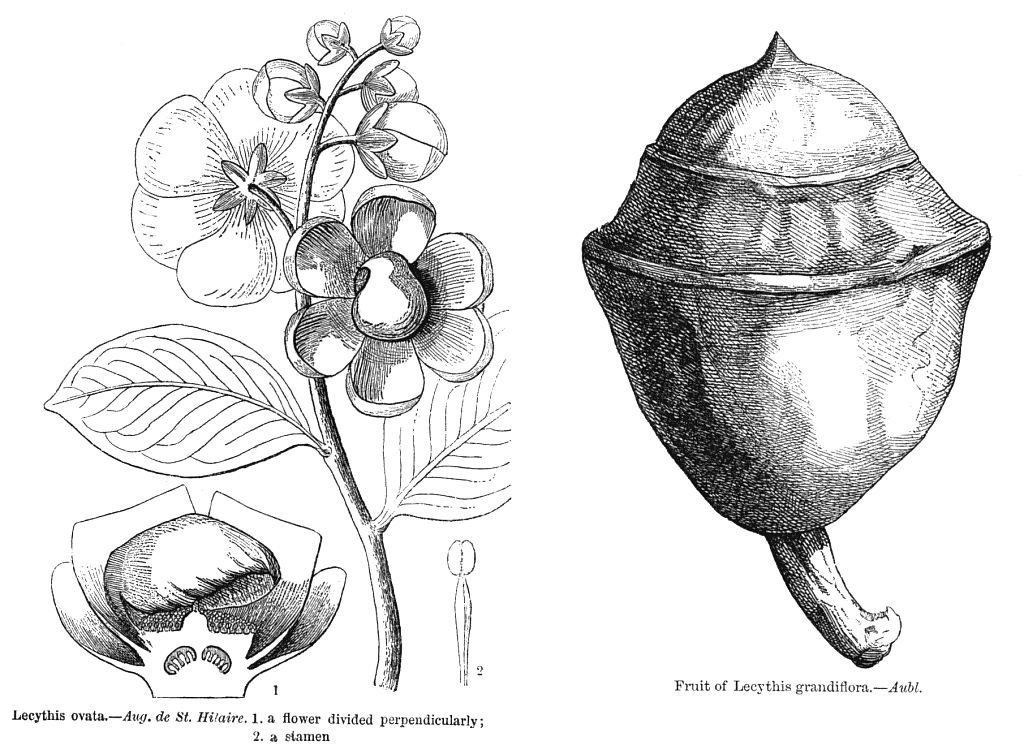
Lecythidaceae PoiteauIncluding GustaviaceaeExcluding Asteranthaceae, Barringtoniaceae, Foetidiaceae, Napoleonaeaceae Habit and leaf form. Trees; without essential oils. Leaves medium-sized, or large; alternate; spiral (aggregated at the tips of the twigs); ‘herbaceous’, or leathery; petiolate; non-sheathing; not gland-dotted; simple. Lamina entire; pinnately veined; cross-venulate. Leaves exstipulate. Domatia occurring in the family (Combretodendron); manifested as pits. General anatomy. Plants with silica bodies. Leaf anatomy. Lamina dorsiventral; without secretory cavities. Stem anatomy. Cork cambium present; initially superficial. Nodes unilacunar. Cortical bundles present (these conventionally orientated). Internal phloem absent. Secondary thickening developing from a conventional cambial ring. The secondary phloem usually stratified into hard (fibrous) and soft (parenchymatous) zones. ‘Included’ phloem absent. Xylem with tracheids (Allantoma), or without tracheids; with fibre tracheids, or without fibre tracheids; with libriform fibres, or without libriform fibres; with vessels. Vessel end-walls simple, or scalariform and simple. Vessels without vestured pits. Wood parenchyma apotracheal, or apotracheal and paratracheal (typically mostly in apotracheal bands). Reproductive type, pollination. Unisexual flowers absent. Plants hermaphrodite. Floral nectaries present. Nectar secretion from the disk. Pollination ornithophilous, or cheiropterophilous. Inflorescence, floral, fruit and seed morphology. Flowers solitary, or aggregated in ‘inflorescences’. The ultimate inflorescence unit (when flowers aggregated) racemose. Flowers regular to very irregular; when irregular, zygomorphic (sometimes the androecium spectacularly so). The floral irregularity involving the perianth and involving the androecium. Flowers cyclic; polycyclic. Free hypanthium absent (‘always with complete fusion of receptacle and ovary’). Perianth with distinct calyx and corolla; 8–12; 2 whorled; isomerous. Calyx usually 4–6; 1 whorled; polysepalous; not persistent (on the fruit); valvate. Corolla 4–6; 1 whorled; polypetalous (usually), or gamopetalous; imbricate; unequal but not bilabiate to regular; deciduous (with the stamens). Androecium 40–1000 (‘many’). Androecial members maturing centrifugally; free of the perianth; coherent (more or less united below, the connate parts sometimes produced on one side of the androecium to form a flat ligule or a hooding structure); 3–5 whorled (? — ‘in several whorls’). Androecium exclusively of fertile stamens, or including staminodes (by abortion of some anthers, in association with one-sided development of the union). Stamens 40–1000 (‘many’); polystemonous; inflexed in bud. Anthers usually versatile; dehiscing via longitudinal slits; tetrasporangiate. Endothecium developing fibrous thickenings. Anther epidermis persistent. Microsporogenesis simultaneous. The initial microspore tetrads tetrahedral, or isobilateral. Anther wall initially with more than one middle layer; of the ‘basic’ type. Tapetum amoeboid. Pollen grains aperturate; 3 aperturate; colporate (or colporoidate, not synocolpate); 2-celled (in Gustavia). Gynoecium 2–6(–10) carpelled. Carpels reduced in number relative to the perianth to increased in number relative to the perianth. The pistil 2–6(–10) celled. Gynoecium syncarpous; synstylovarious, or eu-syncarpous (?); partly inferior, or inferior. Ovary 2–6(–10) locular. Epigynous disk (disks) present (‘usually with an intrastaminal disk, as well as one under C and A’: Airy Shaw 1973). Gynoecium stylate. Styles 1; apical. Placentation basal (Eschweilera), or basal to axile, or axile to apical, or apical. Ovules 1–50 per locule (to ‘many’); pendulous, or horizontal, or ascending; arillate (often, with a funicular aril), or non-arillate; anatropous; bitegmic; tenuinucellate. Outer integument not contributing to the micropyle. Endothelium differentiated. Embryo-sac development Polygonum-type. Antipodal cells formed; 3; not proliferating; ephemeral. Synergids pear-shaped. Endosperm formation nuclear. Fruit often large, fleshy, or non-fleshy; dehiscent, or indehiscent; woody, a capsule, or capsular-indehiscent, or a berry. Capsules when dehiscent, circumscissile (‘monkey pots’). Seeds non-endospermic. Embryo large, rudimentary at the time of seed release to weakly differentiated (as in Bertholletia (the Brazil nut), where it consists mainly of the much thickened hypocotyl), or well differentiated. Cotyledons 2. Seedling. Germination phanerocotylar, or cryptocotylar. Physiology, biochemistry. Cyanogenic, or not cyanogenic. Alkaloids absent (2 species). Iridoids not detected. Proanthocyanidins present, or absent; when present, cyanidin and delphinidin. Flavonols present, or absent; kaempferol, or quercetin. Ellagic acid present (3 genera, 3 species). Saponins/sapogenins present. Sugars transported as sucrose (Lecythis). Geography, cytology. Neotropical. Sub-tropical to tropical. Tropical America. X = 17. Taxonomy. Subclass Dicotyledonae; Tenuinucelli. Dahlgren’s Superorder Theiflorae; Theales. Cronquist’s Subclass Dilleniidae; Lecythidales. APG 3 core angiosperms; core eudicot; Superorder Asteranae; Order Ericales. Species 325. Genera 10; Allontoma, Bertholletia, Cariniana, Corythophora, Couratari, Couroupita, Eschweilera, Grias, Gustavia, Lecythis. Morton et al. (1998) interpret these genera as subfamily Lecythidoideae of their expanded Lecythidaceae. Illustrations. |
Support our developers





{kind=link}
{kind=link}