Germplasm theory
A. Weismann
(1834-1914) demonstrated that pangenesis could not be verified. His popular experiments consisted of cutting the tails of mice and then studying the inheritance. Repeating such a treatment for 22 generations, Weismann found that complete tail structure was still inherited. These experiments of mutilation may appear now rather crude, but results can definitely be used as argument against pangenesis because once the tail was removed, the pangenes or gemmules for the tail will not be available and therefore this structure should not develop in the next generation if pangenesis holds good.
Weismann also proposed his own germplasm theory to account for heredity. According to this theory, the body of an individual can be divided into two types of tissues, germplasm and somatoplasm. The somatoplasm, was not able to enter the sex cells; consequently, the variations
present in the somatoplasm will not be transmitted to next generation. The germplasm, on the other hand, was meant for the reproductive purpose only, so that any change occurring in germplasm will influence the progeny.
Germplasm theory of Weismann was a very significant advancement in our understanding of heredity, since this was for the first time that a distinction between hereditary and environmental variations could be made on a sound basis. However a distinction, between germplasm and somatoplasm in the sense of Weismann may be difficult to make. It is now known that the chromosomes are the main carriers of hereditary characters. This idea was put forward in the form of Chromosome Theory of Inheritance, which will receive a detailed treatment in Physical Basis of Heredity 4. The Chromosome Theory of Inheritance. One may however, note that in recent years hereditary characters have been found in chloroplasts and mitochondria also.
Genotype-phenotype concept, the norm of reaction and developmental noise
In order to make a definite distinction between hereditary and environmental variations, Johannsen in 1909 formulated the genotype-phenotype concept. According to him, the genotype of an individual represents sum total of heredity. On the other hand, phenotype represents features which are produced by interaction between genotype and environment. A genotype can thus exhibit different phenotypes under different conditions. This is referred to as individual's norm of reaction to the environment. Therefore, similar genotypes may not have the same phenotype. Conversely, similar phenotypes do not necessarily mean same genotype. In order to study the interaction of environment and heredity, different methods have been employed.
The first condition for the study of effect of different environments on a genotype would be to have individuals which have same genotype. This can be done by using clones, pure lines or inbred lines. A clone is the progeny of a single plant reproduced asexually e.g. bacterial population derived from a single cell. A pure line is the progeny of a single plant obtained due to continuous self-fertilization. Similarly inbred lines are obtained in cross-fertilizing individuals due to fertilization among closely related lines, such as brothers and sisters. Once clones, pure lines or inbred lines are available, effect of environment can be easily studied, since any observed difference
would be due to environment, the genotype being the same. For a particular genotype, one could tabulate the different phenotypes, resulting due to different environments, in which the genotype grows. Such a tabulation is often done for a particular character, so that only a part of whole genotype is considered. This tabulated set of phenotypes will represent the norm of reaction of the genotype.
 As an interesting case, we may consider the example of himalayan albino rabbits, which are characterized by black colour on feet, ears, nose and tail, the remaining body being white (Multiple Alleles (Based on Classical Concept of Allelomorphism)). If hairs from white part were plucked and the rabbit was allowed to stay at a cold place, the developing hairs were found to be black rather than white. Such changes due to different environments are normally of an adaptive nature, and form the norms of reaction of a genotype.
As an interesting case, we may consider the example of himalayan albino rabbits, which are characterized by black colour on feet, ears, nose and tail, the remaining body being white (Multiple Alleles (Based on Classical Concept of Allelomorphism)). If hairs from white part were plucked and the rabbit was allowed to stay at a cold place, the developing hairs were found to be black rather than white. Such changes due to different environments are normally of an adaptive nature, and form the norms of reaction of a genotype.
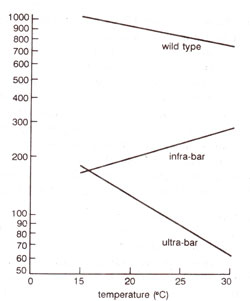
Another interesting example includes the study of the number of facets in the eyes of the following three different mutants grown at temperatures ranging from 0-30°C : (i) wild type, (ii) ultra-bar and (iii) infra-bar. The results of such a study are summarized in Figure 1.2, showing norms of reaction of the three genotypes. Besides the observation that ultra-bar flies are more influenced by variation in temperature, one may notice that higher temperature has opposite effects on ultra-bar (decrease in number of facets) and infra-bar (increase in number of facets) genotypes. One may also notice that although generally the facets in ultra-bar are fewer than in the other two genotypes, but at a particular temperature (16°C), infra-bar may have fewer facets than in ultra-bar. It may, therefore, be necessary to define the environment for a genotype, while discussing its phenotype.
Norms of reaction can also be conveniently studied in vegetatively propagated plants, since cuttings from same plant will give offspring, all
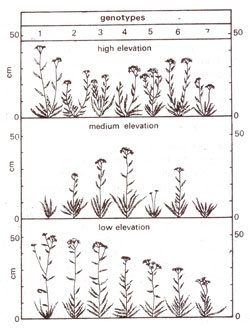
having same genotype. Results of a study of norms of reaction of vegetative offspring of AchUlea, grown at low, medium and high elevations are shown in Figure 1.3. Many plants were collected and three cuttings from each of seven plants, were grown one each at low elevation (30 metres above sea level), medium elevation (1400 metres) and high elevation (3050 metres). Following observations can be made from Figure 1.3. (i) Plants generally grew poorly at medium elevation, (ii) Cuttings from plant 4 grew best at the medium elevation, (iii) Cuttings from plant 1 grew best at low and high elevations, but poorest at medium elevation, (iv) No genotype is superior to all others at all the three elevations, (v) No best genotype or best environment can be selected, because each genotype has its own specific and different norm of reaction.
In case of humans, the norm of reaction can be studied by using zygotic twins (or identical twins), because only these may have similar genotypes for all genes. Since different genes may also interact, we need to have genetically identical (at all loci) plants or animals for the study of the norm of reaction, even when a particular character is being examined. Clones, pure lines and inbred lines, as discussed above, meet this requirement.
In the above discussion, our emphasis has been on the effect of environment in determining the phenotype due to a specific genotype. However, even when environment and genotype are fixed (e.g. wild type at 16°C showing 1000 facets), different individuals or even two eyes of the same individual may differ in the number of facets (ranging from 980 to 1020 in wild type at 16°C). This uncontrolled variation in phenotype may be due to differences in local or internal environment during development, and is described as developmental noise.
Germplasm theory of Weismann was a very significant advancement in our understanding of heredity, since this was for the first time that a distinction between hereditary and environmental variations could be made on a sound basis. However a distinction, between germplasm and somatoplasm in the sense of Weismann may be difficult to make. It is now known that the chromosomes are the main carriers of hereditary characters. This idea was put forward in the form of Chromosome Theory of Inheritance, which will receive a detailed treatment in Physical Basis of Heredity 4. The Chromosome Theory of Inheritance. One may however, note that in recent years hereditary characters have been found in chloroplasts and mitochondria also.
In order to make a definite distinction between hereditary and environmental variations, Johannsen in 1909 formulated the genotype-phenotype concept. According to him, the genotype of an individual represents sum total of heredity. On the other hand, phenotype represents features which are produced by interaction between genotype and environment. A genotype can thus exhibit different phenotypes under different conditions. This is referred to as individual's norm of reaction to the environment. Therefore, similar genotypes may not have the same phenotype. Conversely, similar phenotypes do not necessarily mean same genotype. In order to study the interaction of environment and heredity, different methods have been employed.
Fig. 1.2. Norms of reaction to temperature for three different genotypes for eye size in Drosophila (redrawn from Suzuki ct al., 1986).
Fig. 1.3. Norms of reaction to elevation for seven different AchUlea plants representing seven different genotypes (modified from Carnegie Institute of Washington).
Another interesting example includes the study of the number of facets in the eyes of the following three different mutants grown at temperatures ranging from 0-30°C : (i) wild type, (ii) ultra-bar and (iii) infra-bar. The results of such a study are summarized in Figure 1.2, showing norms of reaction of the three genotypes. Besides the observation that ultra-bar flies are more influenced by variation in temperature, one may notice that higher temperature has opposite effects on ultra-bar (decrease in number of facets) and infra-bar (increase in number of facets) genotypes. One may also notice that although generally the facets in ultra-bar are fewer than in the other two genotypes, but at a particular temperature (16°C), infra-bar may have fewer facets than in ultra-bar. It may, therefore, be necessary to define the environment for a genotype, while discussing its phenotype.
Fig. 1.2. Norms of reaction to temperature for three different genotypes for eye size in Drosophila (redrawn from Suzuki ct al., 1986).
Fig. 1.3. Norms of reaction to elevation for seven different AchUlea plants representing seven different genotypes (modified from Carnegie Institute of Washington).
In the above discussion, our emphasis has been on the effect of environment in determining the phenotype due to a specific genotype. However, even when environment and genotype are fixed (e.g. wild type at 16°C showing 1000 facets), different individuals or even two eyes of the same individual may differ in the number of facets (ranging from 980 to 1020 in wild type at 16°C). This uncontrolled variation in phenotype may be due to differences in local or internal environment during development, and is described as developmental noise.
Support our developers




