The Woody Dicot Stem
First, consider the woody dicot stem. As explained earlier, cells are either meristematic or permanent. Meristematic cellasr e those that retain the ability to divide, whereas permanent cells are those that have lost this ability. Cells at the growing tip of a stem are meristematic. A cluster of young leaves is generally present at the apex of the stem so as to protect the young point. This apex is the terminal bud. Growth in length is accomplished by both divisions of the cells lying under the bud and the elongation of cells lying just below the region of cell division. The cells that continue to divide, called the initials, differentiate into regions, becoming the protoderm, provascular tissue, and ground meristem. The protoderm later becomes epidermis: the provascular tissue becomes primary xylem, primary phloem, and cambium: and the ground meristem produces the remaining primary tissues (that is, the pith, [or cortex] and tissue separating the vascular bundles). mimary tissues are so-called because they are the first formed tissues.As a cell at the apex divides, one of the two resulting cells may lose the ability to divide, thus becoming permanent. The other cell, oriented toward the tip, remains meristematic. In this way, the meristem continually progresses upward.
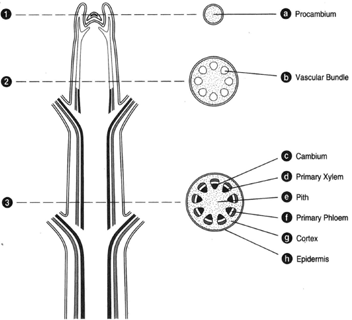 |
| Figure 32-2 At left, longitudinal section of a woody stem tip. At right, corresponding cross sections. (a) Procambium. (b) Vascular bundle. (c) Cambium. (d) Primary xylem. (e) Pith. (f) Primary phloem. (9) Cortex. (h) Epidermis. |
At left of figure 32-2 is a longitudinal section of a stem; at right, corresponding cross sections. No leaf primordia are included. The topmost cross section (figure 32-2a) is cut through a region of apical meristem, where there has been no differentiation of tissues. At the next section down (figure 32-2b), some differentiation of vascular bundles can be seen. The vascular bundles have a primary xylem directed inwardly, a primary phloem directed externally, and a cambium (a zone of meristematic cells), which has not yet produced any divisions. Hence, the only tissues present are primary tissues. In the lowermost section (figure 32-2c), the bundles have coalesced to produce a continuous cylinder of vascular tissue; but there is still no secondary growth. In the center is a zone of undifferentiated (parenchymatous) cells, called the pith. Lying immediately outside of the pith is the vascular cylinder having a primary xylem, a primary phloem, and a cambial layer one cell thick and between the vascular tissues. The cambium has not yet functioned to produce secondary xylem and phloem. Peripheral to the vascular cylinder is another zone of parenchyma (the cortex), and on the outside is the epidermis.
In figure 32-3, a small amount of secondary growth has taken place. While the primary tissues are still detectable, the primary phloem, being pushed outward by the growth of secondary phloem, is becoming crushed and will soon be obscured. All tissues lying outside the cambium tend to be crushed, and the primary tissues in this area are soon entirely obliterated. Enough secondary growth has occurred that the tissues outside the cambium are clearly being destroyed. Primary tissues lying central to the cambium are not subject to crushing and, thus, remain in place through the life of the plant.
Figure 32-4 shows a portion of stem with three years of secondary growth. The primary tissues lying outside the cambium are now entirely obscured, as they have either been sloughed away or replaced by other tissues. Note that as secondary xylem is deposited, the cambium continues to move outward. The cambium, being continuously enlarged, must make on only xylem and phloem, but also additional cambial cells.
The secondary phloem, lying peripheral to the cambium, appears to be compressed and although just as much secondary phloem as secondary xylem is produced, the phloem occupies a comparatively small space (as shown in figure 32-3 of a stem having no more than one year of secondary growth).
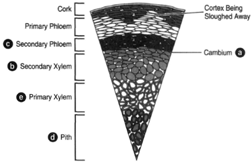 |
| Figure 32-3 Cross section of a woody stem exhibiting no more than one year of secondary growth. (a) The cambium, represented by a single line. (b) The secondary xylem. (c) The secondary phloem. (d) The pith. (e) Primary xylem. |
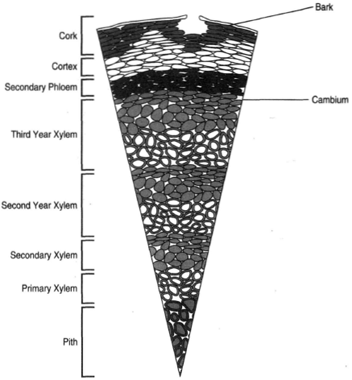 |
| Figure 32-4 Cross section of a woody stem exhibiting three years of secondary growth. |
Figure 32-5 shows a short segment of cambium (at top). When a cambial cell divides (as shown at bottom), one of the cells remains meristematic and the other becomes permanent. If the cell at (1) remains meristematic (and thus remains a part of the cambium), the cell at (2) will be added to the phloem. If the cell at (2) remains meristematic, however, the cell at (1) will become permanent and, thus, part of the xylem. These events alternate (that is, one division yielding a phloem cell, the next division yielding a xylem cell). The cambium also increases itself by producing additional cambial cells.
Secondary xylem is wood. As the wood grows (that is, as additional secondary xylem is produced), recognizable annual rings result. Four kinds of cells are produced in the xylem: vessel elements, tracheids (both of which are involved in conduction), fibers, and parenchyma. The wood formed in the spring tends to be abundant in vessel elements: that formed later in the year tends to be abundant in tracheids. In this manner, recognizable annual rings are formed. A tree one hundred years old will have one hundred annual rings, the oldest of which will be the innermost and in contact with the primary xylem. The newest formed will, of course, be adjacent to the cambium.
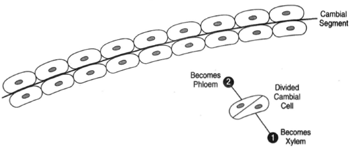 |
| Figure 32-5 At top, a short segment of cambium. At bottom, a cambial cell that has divided. One of the two resulting cells will become permanent; the other will remain meristematic. If cell (2) becomes permanent, it will become part of the phloem. If cell (1) becomes permanent, it will be dead to the xylem. |
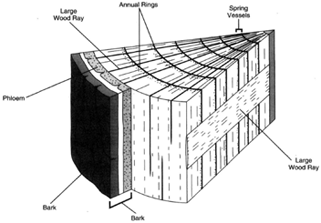 |
| Figure 32-6 Section of wood, showing annual rings. |
The conducting cells of both xylem and phloem are short lived. Only those cells immediately adjacent to the cambium are alive, and only a zone a short distance from the cambium remains functional. The remainder of wood, regardless of tree diameter, is nonfunctional. The center portion of wood, which has lost its water-conducting ability, is called heartwood. Heartwood is often discolored, and its vessels often occluded by incoming seepage of protoplasm from neighboring parenchyma cells. The inflow of cytoplasm causes bladder like formations in the vessel elements, called tyloses (figure 32-7).
A secondary cambium arises outside the phloem and in the remnant of cortex. Here, cells that had lost the ability to divide regain their meristematic ability. This secondary cambium, called the phellogen, produces the outer bark laterally and a greenish layer medially. The latter of these is the phelloderm. Phellogen is also called cork cambium.
Bundles of vascular tissue that pass out into leaves arise from the most recently formed xylem and phloem. Annual additions of xylem and phloem contribute to leaf bundles only once. The strands of vascular tissue that leave the recent zone of xylem and phloem to pass into a leaf are collectively called a leaf trace, and the vacant space immediately above the leaf trace and in the vascular cylinder is called a leaf gap (figure 32-8).
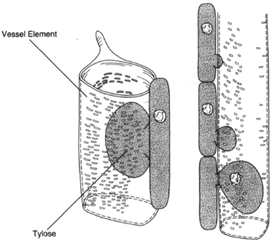 |
| Figure 32-7 Tyloses result from an inflow of cytoplasm from neighboring parenchyma cells to the vessel elements. Tyloses tends to top the flow through vessels. |
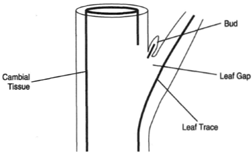 |
| Figure 32-8 The cambial tissue and the cells immediately adjacent to it. The leaf trace is the vascular tissue that leaves the main cylinder. The leaf gap is the pace left above the leaf trace. |
The most prominent elements of phloem are those that compose the sieve tubes. These elements are elongated cells arranged end to end and having perforated end walls. The end walls are called sieve plates. At the side of each sieve cell is a parenchymatous cell called the companion cell. There appears to be living cytoplasm in the sieve-tube element, although the mature cell does not have a nucleus. Strands of cytoplasm run through the holes of the sieve plates to neighboring sieve-tube elements.
The sieve-tube elements, having cytoplasm, are living. In xylem, substances move through dead cells. The presence of cytoplasm is essential to the function of phloem cells. The flow of solutions through sieve tubes is hypothesized to result from pressure; the leaves develop a higher turgor pressure and push solutions through the phloem.
Phloem exudate contains 5 to 20 percent sugars, proteins, minerals, and hormones. Many substances can move through phlooenmly when combined with sugars. While the direction of fluid movement in xylem is primarily upward, fluids move both upward and downward through phloem.
Deposits of callose (a polysaccharide of glucose units) form at the sieve plates. In time, these deposits clog the holes in the sieve. plates. Sieve-tube elements and their corresponding companion cells are daughters of a common cell. A sieve-tube mother cell undergoes division to produce the sievetube element and the companion cell that resides beside it.
In addition to the sieve tubes and companion cells, parenchyma and fibers are also present. The fibers may be impregnated with lignin.
The cells lying outside of the phellogen, the outer bark, become suberized and soon die. Suberin is a waxy material that makes the outer bark resistant to invasion of bacteria and mildew and retards the loss of water. Bark often possesses openings called lenticels, which are believed to allow an exchange of gases (although this has not been proven). A number of plant species, including woody plants, do not have lenticels.
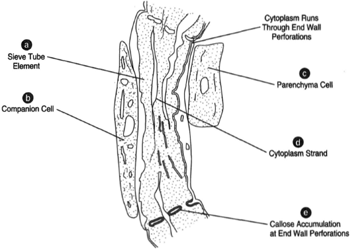 |
| Figure 32-9 (a) Sieve-tube element, (b) Companion cell, (c) Parenchyma cell, (d) Cytoplasm of sieve cell, (e) Callose accumulation. |
 |
| Figure 32-10 A lenticel, an eruption in the surface of the bark. |
Support our developers




