The Evolution of Eukaryotic Cells
At one time, prokaryotes were thought to be more closely related to a postulated progenote (the common ancestor of all cells, before there was a genome) than were eukaryotes, and all prokaryotes were thought to be more closely related to one another than to any eukaryote. Most prokaryotic species can be further classified as eubacteria. The other prokaryotic subkingdom, archae, occupies the kinds of environments that were presumed to be widespread when life first evolved. Hence, it was commonly believed that eubacteria evolved from primitive archae and eukaryotes evolved from eubacteria. Gradually, however, many more differences were found to separate the two subkingdoms. Some archae traits are shared with the eubacteria (they are both prokaryotes), whereas others are shared with eukaryotes (e.g., genes for rRNAs and tRNAs contain introns). Based on his analysis of nucleotide sequences in the highly conserved 16S rRNAs from many organisms, Carl Woese proposed in 1977 that archae are as different from eubacteria as either group is from the eukaryotes. Today, it is thought that all three lines have descended from the same progenote.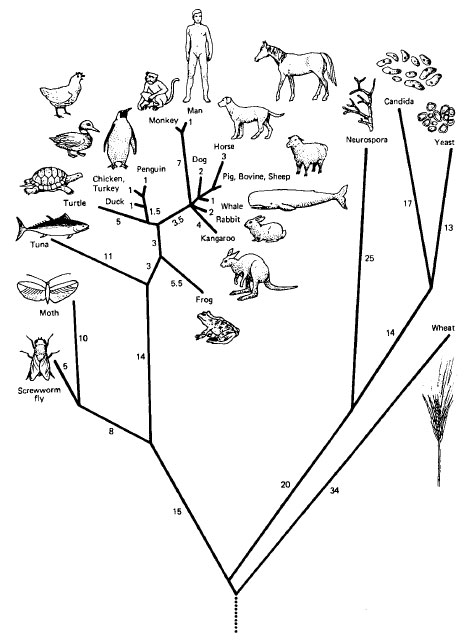 |
| Figure 12-1 A phylogenetic tree based on homologies between cytochrome c molecules in various organisms. Branch length is represented by the most likely number of point mutations that occurred during evolution of these species. |
Organisms with a nucleus may have evolved as long ago as 3.5 billion years, but how the first nuclear membrane arose remains a mystery. According to the membrane proliferation hypothesis, one or more invaginations of the plasma membrane in the progenote coalesced internally to surround the genome, became severed from the plasma membrane, and formed a double-layered nuclear membrane. The manner of infolding of the plasma membrane shown in Figure 12-2 accounts for the fact that the nucleus of modern eukaryotic cells is enclosed within a "double membrane" consisting of two lipid bilayers. Note that a portion of the ER is continuous with the outer membrane of the nuclear envelope.
The origin of mitochondria in younger eukaryotes may be explained by the endosymbiotic theory. Some ancient cells were capable of ingesting food particles by endocytic invaginations of their plasma membranes. It is possible that at least one large, fermenting, feeder cell engulfed one or more smaller respiratory bacteria, but failed to digest them. This endosymbiont was able to survive in an environment where nutrients were abundant and it could hide from other predatory cells. In turn, the host feeder cell gained the energetic advantages of oxidative respiration over fermentation. These complementary advantages evolved into a symbiotic ("living together") relationship wherein neither entity can survive without the other. Part of this mutual adaptation involved the transfer of most of the genes of the bacterial endosymbiont into the nucleus of the host cell. Most negatively charged molecules, including mRNAs, tRNAs, rRNAs, and some proteins, that cannot cross the membrane of these organelles must still be encoded by the genomes of these organelles. This process is proposed to have given rise to the mitochondria of modern eukaryotic cells at least 1.5 billion years ago.
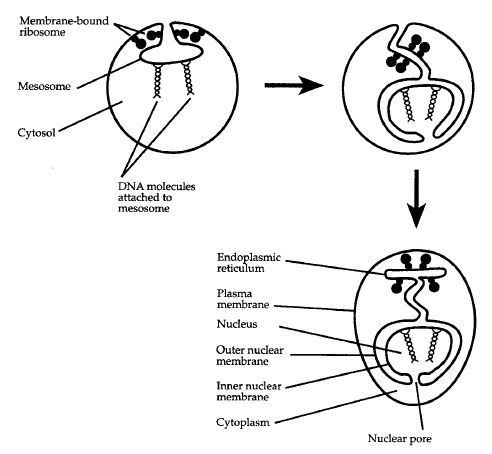 |
| Figure 12-2 Formation of a double nuclear membrane. |
Astronger case can be made for the evolution of chloroplasts by endosymbiosis than that for mitochondria. An aerobic, eukaryotic feeder cell (one that had already evolved mitochondria) is proposed to have engulfed one or more eubacteria (related to cyanobacteria) that were capable of oxygenic photosynthesis. In the process of evolving into chloroplasts, the endosymbionts relinquished some of their genes to the nuclear genome, but not as many as did the endosymbionts that evolved into mitochondria. Like the mitochondria, the protochloroplasts had to retain all of the genes specifying tRNAs and rRNAs for protein synthesis within the chloroplast.
Much evidence supports the endosymbiotic theory for the origin of chloroplasts and mitochondria. These organelles are approximately the same size as bacteria. The genomes reside within a single, circular DNA molecule that is devoid of histone proteins, like bacteria. Both organelles reproduce asexually by growth and division of existing organelles in a manner similar to binary fission. Protein synthesis in mitochondria and chloroplasts is inhibited by a variety of antibiotics that inactivate bacterial ribosomes, but have little effect on cytoplasmic eukaryotic ribosomes. Nascent polypeptides in bacteria, mitochondria, and chloroplasts have N-formylmethionine at their amino ends. Mitochondria and chloroplast genomes encode the tRNA and rRNA molecules for their own protein- synthesizing systems. The ribosomes in both organelles resemble bacterial ribosomes in size and structure. Lastly, the endosymbiotic theory accounts for the fact that both organelles have double membranes. The inner membrane corresponds to the plasma membrane of the ancestral endosymbiont; the outer membrane represents the plasma membrane of the ancestral feeder host cell.
Notes
A type of purple, photosynthetic bacteria that had lost its photosynthetic ability and retained its respiratory chain is hypothesized to represent the endocytosed bacteria.
One theory suggests that the flagella and cilia of eukaryotes originated from motile, symbiotic bacteria on the surface of ancestral eukaryotic cells.
Support our developers




