Inheritance of characteristics
Content
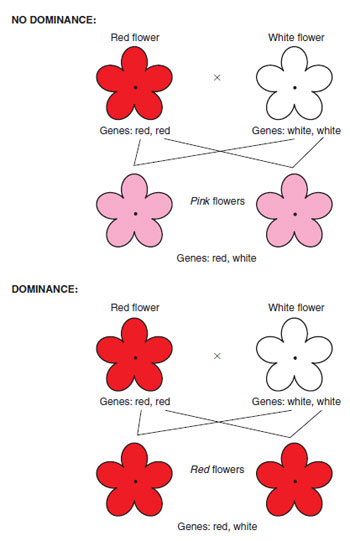
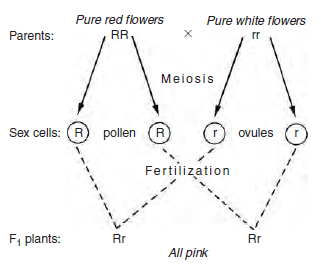
As mentioned above, genetic information is passed from parent to offspring when material from male and female parent comes together by fusion of the sex cells. Genes from each parent can, in combination, produce an intermediate form, a mixture of the parents’ characteristics in the offspring; e.g. a gene for red flowers inherited from the male parent, combined with a gene for white flowers from the female parent, could produce pink-flowered offspring, if both conditions are equal (see below). If one gene, however, was completely dominant over the other, e.g. if the red gene inherited from the male parent was dominant over the white female gene, all offspring would produce red flowers (see Figure 10.6). The non-dominant (recessive) white gene will still be present as part of the genetic make-up of the offspring cells and can be passed on to the next generation. If it then were to combine at fertilization with another white gene, the offspring would be white-flowered. The example above considers the inheritance of one pair of genes by a single offspring (monohybrid cross) . However, many sex cells are produced from one flower in the form of pollen grains and ovules, and these give rise to many seeds in the next generation. The plant breeder tries to predict the ratio (or percentage) of offspring with each option (in this case, the ratio of red- flowered offspring to white-flowered offspring).To understand this prediction process, consider now the same example of flower colour in a little more detail. If a red-flowered plant, containing two genes for red (described as pure), is carefully fertilized (crossed) with a ‘pure’ white-flowered plant, the red-flowered plant supplies in this case pollen as the male parent, and the white-flowered plant supplies ovule as the female parent. As both parents are pure, the male parent can produce only one type of sex cell, containing the ‘red’ version (allele) of the gene, and the female parent only the white (allele). Since all pollen grains will carry ‘red’ genes and all ovules ‘white’, then in the absence of dominance, the only possible combination for the first generation (or F1) is pink offspring, each containing an allele for red and an allele for white (i.e. impure). Figure 10.7 illustrates this inheritance by using letters to describe genes, R to represent a red-inducing gene, and r to represent a white-inducing gene.
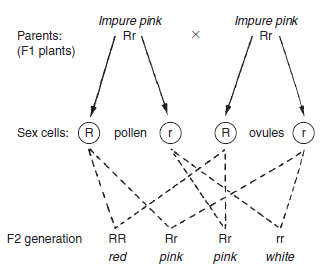
The genotype (the genetic make-up of a cell) can be represented by using letters, e.g. Rr. The phenotype (the outward appearance e.g. red, pink or white flower) results from the genotype’s action. If these plants from the F1 generation were now used as parents and crossed (or perhaps selfpollinated), then the results in the second or F2 generation would be as shown in Figure 10.8, i.e. 25 per cent of the population would have the phenotype of red flowers, 50 per cent pink flowers and 25 per cent white flowers (a ratio of 1:2:1). The plant breeder, by analysing the ratios of each colour, would be able to calculate which colour genes were present in the parents, and whether the ‘colour’ was pure. More usually, one of the alleles at the gene site exhibits dominance and the other is recessive. As a result, when offspring inherit both alleles (dominant and recessive), there is no intermediate phenotype; only the dominant gene expresses itself (the recessive is masked). It was the monk Gregor Mendel working in a monastery in what is now Brno in the Czech Republic in the middle of the eighteenth century who laid the foundations of genetics with his breeding experiments. He used peas and worked with single characteristics such as the height of the plant. He crossed tall with short plants and found that in the second or F2 generation the plants were either tall or short; there were no middlesized phenotypes. They were found to be in the ratio of three tall to one short plant. Similarly, he crossed pure (homozygous) round pea parents (RR) with pure (homozygous) wrinkled pea parents (rr). When crossing the two pure parents (RR × rr), the genotype of the first generation, or F1, follows the pattern as shown in Figure 10.7, but all the plants produced round peas, i.e. the round pea allele was dominant in peas. When the second or F2 generation was produced the genotype followed the pattern shown in Figure 10.8, but there were three round peas to every one wrinkled pea type. Both RR and Rr genotypes produced the same phenotype; only a double recessive (rr) produced plants with wrinkled peas. He went on to look at other simple ‘single gene characteristics’ including seed colour (yellow dominant and green recessive) and flower colour (purple dominant and white recessive). In all these cases he found that the ratio of phenotypes in the second or F2 generation was 3:1. By observation he established the Principle of Segregation which states that the phenotype is determined by the pair of alleles in the genotype and only one allele of the gene pair can be present in a single gamete (i.e. passed on by a parent).
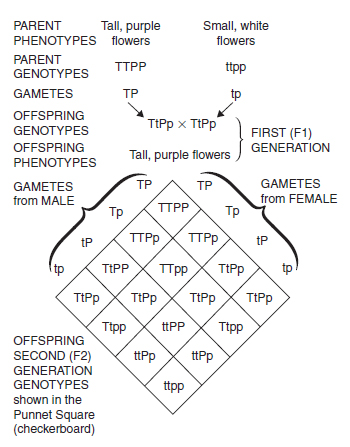
Dihybrid cross. Mendel then went on to investigate the crossing of plants that differed in two contrasting characters. The results of crossing tall purple-flowered peas with short white-flowered ones produced all tall purple-flowered peas. As Mendel showed, they all had the same genotype (TtPp) and the same phenotype. The F2 generation produced from these parents is illustrated in Figure 10.9 , where the combinations are shown in a Punnet Square (or ‘checkerboard’ – a useful way of showing the genotypes produced in a cross). Note that each gene has behaved independently; there are 12 tall to four short (ratio 3:1) and 12 purple to four white-flowered plants (3:1). This example illustrates Mendel’s Principle of Independent Assortment which states that the alleles of unlinked genes (i.e. genes not on the same chromosome) behave independently at meiosis. The cross involving two independent genes produces combinations of phenotypes in the ratio 9:3:3:1. For the cross TtPp × TtPp, the genotypes produced are shown in Figure 10.9 and the phenotypes are as follows:
Not all genes act independently in the way shown above. Some are linked by being on the same chromosome and so they will usually appear together. The number of grouped genes is equal to the number of chromosome pairs for the species (seven for peas).
Breeders may choose to produce seeds for the commercial market which are all F1 offspring or sometimes called F1 hybrids. F1 hybrid seeds are important to the grower since, given a uniform environment, all plants of the same cultivar will produce a uniform crop because they are all genetically identical (see Figure 10.10). Crops grown from F1 hybrid seed such as cabbage, Brussels sprouts and carrots can be harvested at one time and they have similar characteristics of yield. Similarly, F1 hybrid flower crops will have uniformity of colour and flower size. Another feature of F1 hybrids is hybrid vigour. Plants crossed from parents with quite different characteristics will display the feature to a marked extent, giving outstanding growth, especially in good growing conditions. The desirable characteristics of the two parents, such as disease resistance, good plant habit, high yield and good fruit or flower quality, may be incorporated along with established characteristics of successful commercial cultivars by means of the F1 hybrid breeding programme. F1 hybrid seed production first requires suitable parent stock, which must be pure for all characteristics. In this way, genetically identical offspring are produced, as described in Figure 10.7. The production of pure parent plants involves repeated self pollination (selfing) and selection, over eight to twelve generations, resulting in suitable inbred parent lines. During this and other self pollination programmes, vigour is lost (the parent plants do not look impressive) but, of course, the vigour is restored by hybridization. The parent lines must now be cross-pollinated to produce the F1 hybrid seed. It is essential to avoid self pollination at this stage, therefore one of the lines is designated the male parent to supply pollen. The anthers in the flowers of the other line, the female parent, are removed, or treated to prevent the production of viable pollen. The growing area must be isolated to exclude foreign pollen, and seed is collected only from the female parent. This seed is more expensive than most other commercial seed, due to the complex breeding programme requiring intensive labour. Seed collected from the planted commercial F1 hybrid crop represents the F2, and will produce plants with very diverse characteristics (Figure 10.8). Some F2 seed, however, is deliberately produced by breeders for flowering plants, such as geraniums and fuchsias, where a variety of colour and habit is required for bedding plant display. |

Support our developers




