Glaucophyta
As already described in previous sections these algae possess inclusion termed cyanelles that are probably symbiotic cyanobacteria functioning as chloroplasts. Each cyanelle, surrounded by a reduced peptoglycan cell wall (except in Glaucosphaera sp.), is enclosed in a vesicle of the host cytoplasm.
Cyanelles do not fix molecular nitrogen in contrast with cyanobacteria; they contain polyphospate
granules and a conspicuous central carboxisome similar to the pyrenoids of other algae. The
thylakoid are not stacked but they are single and equidistant with a concentric arrangement.
Cyanelle pigments are chlorophyll
a and β-carotene, which represents the main carotenoid.
Interthylakoidal phycobilisomes contain allophycocyanin and phycocianin. Phycoerytrin is
absent from Glaucophyta but phycoerythrocyanin can be found in some species.
Rhodophyta

FIGURE 2.77 Transmission electron microscopy image of rhodophyte thylakoid showing with regularly arranged phycobilisomes. (Bar: 0.05 µm.)
Ultrastructurally, red algal chloroplasts are composed of a double-membrane envelope inside of
which are one or more parallel, thylakoidal photosynthetic lamellae. These chloroplasts are not
associated with the endoplasmic reticulum, a feature shared with Glaucophyta and Chlorophyta.
Encircling thylakoids are present in all Florideophyceae and in some taxa of Bangiophyceae
while the thylakoids in the other red algae lie equidistant and are single, that is, not stacked,
unlike any other group of eukaryotic algae (except Glaucophyta), and typically oriented parallel
to each other. All thylakoids have phycobilisomes attached to their stromal surface, which
contain the accessory phycobiliprotein pigments, that is, allophycocianin, phycocyanin, and five
forms of phycoerythrin (Figure 2.77). Chlorophyll a is the only chlorophyll present in the thylakoid
membrane, together with carotenoids such as β-carotene and lutein. Plastid number, shape, and
position (many, discoid, and parietal) is rather uniform throughout the Florideophyceae, and
pyrenoids may or may not be present. A single stellate plastid with a central pyrenoid is commonly
associated with bangiophycidean red algae, such as
Phorphyridium. DNA is organized into numerous nucleoids scattered throughout the chloroplast.
FIGURE 2.77 Transmission electron microscopy image of rhodophyte thylakoid showing with regularly arranged phycobilisomes. (Bar: 0.05 µm.)
HeterokontophytaSome ultrastructural features of the chloroplast compartments of these algae are common to all the
seven classes of the division, with few exceptions. Four membranes surround the chloroplasts, the
outer two being the chloroplast endoplasmic reticulum and the inner two being the chloroplast envelope. When the chloroplasts are located close to the nucleus, the chloroplast endoplasmic
reticulum is continuous with the nuclear envelope. In the Xanthophyceae, this connection is not
the rule. Thylakoids are grouped into lamellae of three, which are two in some Raphidophyceae,
with varying degrees of coherence depending on the species. Thylakoids from adjacent lamellae frequently interconnect across the stroma. In all the classes, with the exception of Eustigmatophyceae
and
Chattonella (Raphidophyceae), one lamella runs around the periphery of the chloroplast beneath the chloroplast envelope, enclosing all the other lamellae. The lamella is called girdle lamella.
One or more plate-like (Chrysophyceae, Figure 2.78a and 2.78b; Eustigmatophyceae,
Figure 2.79a and b), discoid (Xanthophyceae, Raphidophyceae, and Dictyochophyceae) or
ribbon-like (Phaeophyceae), plastids are typically present, often lobed, parietal or located in
close connection with the nucleus. In the Bacillariophyceae, chloroplasts are the most conspicuous
feature, and their number and shape are consistent features of taxonomic importance. They may be
rounded or lobed discs or large plate-like with or without lobed margins, and may range from one to
two, four or more. A typical centric diatom has many disc-shaped plastids, arranged close to the
periphery surrounding a large central vacuole or scattered throughout the cell. The raphid
diatoms tend to have large chloroplasts (one to four) lying along the girdle with central nucleus
flanked by two large vacuoles.
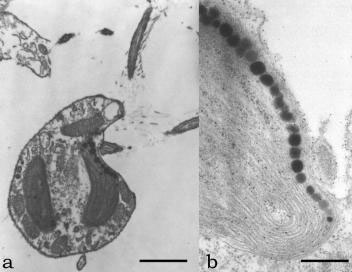
FIGURE 2.78 Transmission electron microscopy image of O. danica in longitudinal section, showing the chloroplast (a) (Bar: 3 mm). Transmission electron microscopy image of a chloroplast at higher magnification showing the thylakoid membrane and the eyespot globules (b). (Bar: 1 µm.)
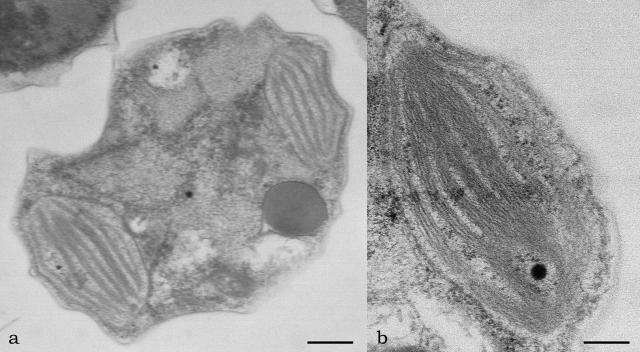
FIGURE 2.79 Transmission electron microscopy image of Nannochloropsis sp. in transverse section, showing the chloroplast (a) (Bar: 0.50 mm); chloroplast at higher magnification (b) (Bar: 0.10 µm).
The chloroplast DNA is ring-shaped and located in the region between the girdle lamella and
the others in all the classes, with the exception of the Eustigmatophyceae, where it is organized into
many dot-like nucleoids, which may be united in a sort of reticulum. The main photosynthetic
pigment is chlorophyll
a, which is the only chlorophyll present in the Eustigmatophyceae.
In addition, chlorophylls of the c group occur, both c
1 and c
2 (Chrysophyceae, Rhaphidophyceae, Phaeophyceae, Dictyocophyceae, and in only extremely low concentrations in Xanthophyceae) or only c
2 (Bacillariophyceae). The most important accessory pigment is fucoxanthin in Chrysophyceae, Bacillariophyceae, Dictyocophyceae, and Phaeophyceae, violaxanthin in Eustigmatophyceae, and vaucheriaxanthin in Xanthophyceae. Other accessory pigments are β-carotene and xanthophylls such as diadinoxanthin, heteroxanthin, vaucheriaxanthin, antheraxanthin, and lutein. In the Raphidophyceae marine and freshwater species differ in their accessory pigments, marine
species possessing mainly β-carotene, fucoxanthin, and violaxanthin, and freshwater species
having β-carotene, diadinoxanthin, heteroxanthin, and vaucheriaxanthin.
Pyrenoids are present in all the classes, except in the zoospores of the Eustigmatophyceae and
in the freshwater species of Raphidophyceae. They are of a semi-immersed type, attached to the inner face of the chloroplast, pear-shaped in the Phaeophyceae, or stalked in the Eustigmatophyceae.
They can be one or many (Bacillariophyceae and Phaeophyceae). No storage material or
capping vesicles have been found to be associated with pyrenoids, but lipid or oil droplets normally
distributed randomly in the chloroplast matrix are often concentrated at the periphery of the
pyrenoid.




