Techniques leading to nucleosome discovery
X-ray diffraction and electron microscopy
While studying X-ray diffraction patterns of chromatin, M.F. Wilkins and V. Luzzati in the year 1960 observed patterns which suggested repeating units in chromatin. Similar repeats were inferred from X-ray diffraction of a mixture of DNA and histones also. Electron microscopy was also used for a study of chromatin structure. In 1974, A.L. Olins and D.E. Olins reported results of electron microscopy of chromatin obtained from interphase nuclei lysed in water. Under electron microscope, chromatin fibre appeared as arrays of spherical particles, about 10 nm (1 nm = 10-9 m = 10 Å) in diameter, connected by filaments, which were about 2 nm in diameter. These particles were called v (nu) bodies.
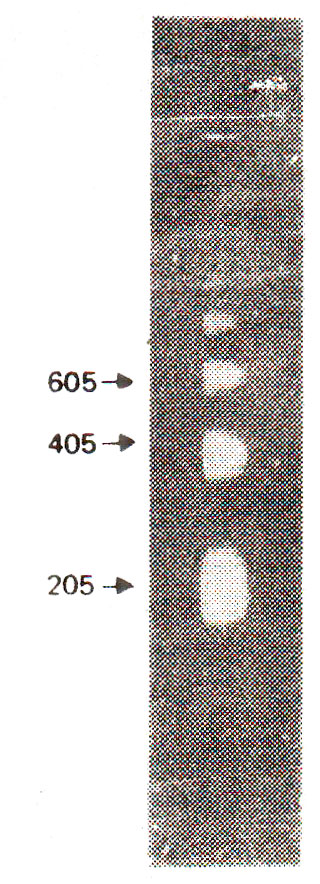
Fig. 27.1. Multimeric series of DNA bands after nuclease digestion of chromatin followed by gel electrophoresis.
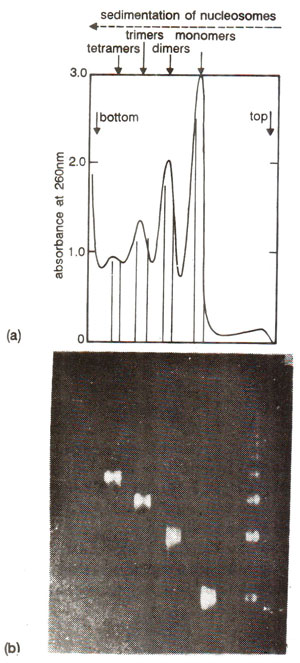
Fig. 27.2. (a) Results of sedimentation of nucleosome preparation on sucrose gradient showing separation of monomers, dimers, trimers and tetramers. (b) Gel electrophoresis of DNA purified from fractions in (a) above suggesting that each multimer had a multiple of unit DNA length, (redrawn from Lewin's 'Genes IV).
X-ray diffraction patterns of native chromatin could be compared with those of chromatin, reconstituted from DNA and histones to find out the degree of successful reconstitution. It was shown by T. Kornberg (T. KornfTerg is different from A. Kornberg of DNA replication fame), that 90% reconstitution was achieved, when DNA was mixed with unfractionated histone preparation. However, when purified histones were used, reconstitution failed suggesting that some denaturation of histones takes place during purification, which destroys the compactly folded native conformation of chromatin.
Nuclease digestion and regular distribution of histones
Nucleases are a class of enzymes which will cause fragmentation of DNA at those sites which are not protected by proteins like histones. Therefore, it was expected that through a study of DNA fragments, location of these sites of naked DNA will be possible and information will thus be available about the nature of association of histones with DNA. This technique of nuclease digestion was utilized in 1973 by Hewish and Burgoyne for fragmentation of chromatin of rat liver cells. The fragments were separated by gel electrophoresis. It was found that fragments of different sizes were produced which formed distinct bands representing sizes which were multiples (two, three, four times, etc.) of size of the smallest unit. This information came from further detailed study of bands (Figs. 27.1, 27.2). On this other hand, when DNA alone and not the chromatin, was fragmented with same nuclease enzyme, uniform smear father than bands were observed on gel. Appearance of uniform smear rather than bands meant that fragments obtained due to nuclease digestion of naked DNA, gave a gradient of size, rather than being multiples of smallest unit size as shown through distinct bands in nuclease digestion of chromatin. These studies of rat liver chromatin clearly demonstrated that rat nuclease cleaved chromatin at sites spaced at regular intervals along the DNA, while naked DNA was cleaved at random. Therefore, it was concluded that histones confer a regular pattern of protection against nuclease and, that the proteins should be distributed along the DNA in a regular periodicity.
Fig. 27.1. Multimeric series of DNA bands after nuclease digestion of chromatin followed by gel electrophoresis.
Fig. 27.2. (a) Results of sedimentation of nucleosome preparation on sucrose gradient showing separation of monomers, dimers, trimers and tetramers. (b) Gel electrophoresis of DNA purified from fractions in (a) above suggesting that each multimer had a multiple of unit DNA length, (redrawn from Lewin's 'Genes IV).
There are certain chemicals which help in chemical cross linking and thus facilitate in the study of cross linking between molecules, if any. When this technique was applied to histones in solution, it was discovered that H3 and H4 were present as a tetramer (or a pair of dimers) i.e. (H3)2.(H4)2 and not as a simple dimer (H3.H4). This tetramer nature was subsequently confirmed in chromatin also. The relationship of this tetramer with other histones and DNA was also studied. It was shown that chromosomes of most organisms had equal number of molecules of histones H2A, H2B, H3 and H4. It was also shown that 25 nucleotides of DNA were present, per histone molecule. Since H3 and H4 are present as tetramers, (H3)2.(H4)2, each repeating unit may have one such tetramer (described as kernel) and two molecules each of H2A and H2B (because in chromatin H2A = H2B = H3 = H4). Histones will thus form an octamer, (H3)2.(H4)2/(H2A.H2B)2 associated with 200 bases in each repeating unit. One molecule of H1 is also associated with each octamer in a nucleosome unit. However, H1 was not linked with other histones, so that in core particle H1 was absent.
Support our developers




