Applying Genetics
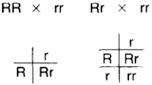 |
| Figure 5-8 A test cross (or back cross) can be used to determine genotype. Here two, round- seeded forms are crossed with the homozygous recessive to reveal the genotype. Round-seeded offspring indicate that the round-seeded parent was homozygous; any wrinkled-seeded offspring indicate that the round-seeded parent was heterozygous. |
Now consider a problem. We have some round-seeded peas. The phenotype is visible, but the genotype is unknown. The peas could be homozygous (RR), or they could be heterozygous (Rr). How can we determine the genotype? This is done by a test cross, also known as a back cross. A test cross is crossing an unknown genotype with a homozygous recessive. If all progeny of such a cross display the dominant traits, the unknown genotype is homozygous. If, however, the recessive trait should appear among even only a few of the progeny, the unknown genotype is heterozygous.
Consider a back cross between turnips. In turnips, purple (P) is dominant and red (p) is recessive, and cylindrical (C) is dominant and spherical (c) is recessive. When a purple- cylindrical turnip is crossed with a red-spherical turnip, the F1, as expected, has both dominant
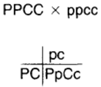 |
| Figure 5-9 A homozygous, purple, cylindrical turnip is crossed with a homozygous, red, spherical turnip |
traits (see figure 5-9). Now, use the members of the F1 in a back cross and follow the procedure of recording only the kinds of gametes that can be produced. Here, the female parent can produce four kinds of gametes, the male only one. At the top of the column in figure 5-10, the progeny show both dominant traits: purple and cylindrical. The second in the column shows dominant color and recessive shape: purple and spherical. The third in the column shows recessive color and dominant shape: red and cylindrical. The fourth in the column shows both recessive traits: red and spherical. According to the principle of independent assortment, the expected ratio would be 1: 1: 1: 1. Yet, a ratio of 7: 1: 1:7 was actually observed. How could this be? The answer relates to the traits being on the same chromosomes and, thus, being linked. Based on this knowledge, one might expect there to be only two kinds of gametes: PC and pc. If linkage were unalterable, this would be the case. But as described in chapter 3 on mitosis and meiosis, crossing over allows linkage alterations; crossing over, then, is what makes possible the combinations PC and PC.
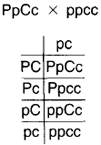 |
| Figure 5-10 The progeny of the cross in figure 5-9 are subjected to a test cross, being crossed with the pure recessive for both traits. |
Having examined several examples illustrating the law of dominance, where a gene finds full expression and its homologue is suppressed, consider now a situation wherein both genes of a pair are equally effective in their influence. Mirubilis jalapa is an annual plant often planted in flower gardens. (Its common name is "Four O'clock' because its flowers are said to open at approximately that time of day.) If a pure red variety (RR) were crossed with a pure white form (rr), the heterozygous offspring would have pink flowers, the influence of both sets of genes being expressed. This is an example of incomplete dominance. If such pink flowers are self pollinated or crossed with other pink forms, the offspring will be in the ratio 1:2:1: one-fourth red, two-fourths pink, and one-fourth white.
Dominance can be affected by environmental conditions. If Chinese primroses are grown at 68°F, for example, they will produce red flowers; but if grown at 80°E white flowers will result.
Following is an example of multiple genes affecting a single trait. There are four genes (two pairs) that influence grain color in wheat. When red and white-grained varieties are crossed, medium-red offspring result. When these offspring are interbred, the grain of the next generation ranges in color from red to white. This is called polygenic inheritance. (See figure 5-11).
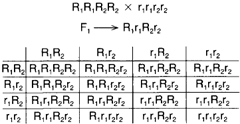 |
| Figure 5-11 Polygenic inheritance. If four Rs are present in the progeny, dark grains will be noted; if three Rs are present, the grain color will be medium-dark. If two Rs are present, grain color will be medium red; and if no Rs are present (that is, only small rs are present), the grain color will be white. |
Earlier, Hugo DeVries' work with Oenothera was mentioned. DeVries observed that from time to time, a progeny displayed a trait that seemed to come from neither parent. He proposed that such a condition resulted from an unexpected change in a gene. He called this mutation. Navel oranges, nectarines, numerous types of potatoes, and variegated shrubs all result from mutation. Asexual means of propagation of such forms makes it possible to retain desired features.
Many changes occur for reasons other than mutation. (In fact, approximately only one in one thousand changes can be traced to mutations.) Other factors which affect phenotype are alterations in chromosomal configuration such as deletion of parts of chromosomes or duplication of chromosomal segments. Changes in the number of chromosomes also alter the phenotype. Most mutations produce no benefit for organisms and even have harmful consequences, but a few are beneficial. Tens of thousands of plants may be treated in the hope of causing a desired change.
Colchicum autumnale, the autumn crocus, produces the alkaloid colchicine, which can produce tetraploidy (4N)y arresting some part of the mitosis process. Extract from this plant is used in extreme dilution. When dividing plant cells are placed in contact with a colchicine solution, mitosis is arrested in late prophase, after chromosome replication has taken place. The cells can then be removed from the solution, at which time they recover and start the process of mitosis again from the beginning. The result is that the chromosomes replicate twice, doubling the chromosome number (tetraploidy). There are clear benefits to using the autumn crocus for this purpose. Polyploid plants are usually more vigorous than are diploid forms, and they sometimes produce larger fruits. Most cultivated irises and many lilies are tetraploids. Baldwin apples, pink beauty tulips, and Japanese flowering cherries are triploids.
Finally, although we have focused on situations wherein one gene affects one trait, there are situations wherein one gene affects more than one trait. This is called pleiotropy. For example, the gene involved in the synthesis of lignin may also cause numerous other effects.
Support our developers




