Coniferales
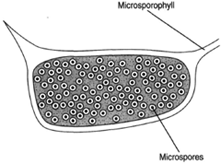 |
| Figure 27-1 Microsporangium with microspores. The microsporangium is borne on the lower surface of the microsporophyll. |
Consider the pine as an example of Coniferales. Pines are monoecious, producing both male and female cones (strobili) on the same tree. Thus, they produce both microspores and megaspores. Spore formation begins when the trees are approximately ten years old. The male cones, perhaps one-half inch long, are borne in clusters on side branches. The scales of the cone are microsporophylls, and two microsporangia are found on the underside of each scale. Microspore mother cells in the microsporangium undergo meiosis to produce haploid microspores. The microspore is an immature pollen grain with a two-layered wall: an outer exine and an inner intine. The exine bulges away from the intine to produce air sacs, which aids wind dispersal of the pollen. Several nuclear divisions occur before the pollen is mature and released. These divisions produce two prothallial cells, a generative cell, and a tube cell. At this stage, the microsporangium splits open to release the pollen grains.
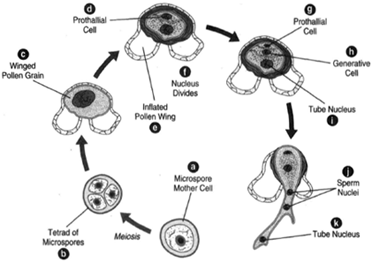 |
| Figure 27-2 Development of the male gametophyte from a microspore. (a) Microspore mother cell. (b) Microspore mothecre ll undergoes meiosis to produce a tetraodf microspores. (Only three microspores are shown here.) (c) Winged pollen grain. (d) The prothallial cell. (e) The inflated wing of the pollen. (f) The nucleus divides. (g) The prothallial cell. (h) The generative cell. (i) The tube nucleus. The generative cell divides to form a stalk cell and a body cell. (j) The body cell then divides to form two sperm nuclei. It is then a mature male gametophyte. (k) The tube nucleus. |
In the female cones, megasporangia are on the upper surfaces of the megasporophylls. In each megasporangium, ma egaspore mother cell undergoes meiosis to produce a tetrad of megaspores. Three of these megaspores degenerate and disappear. The remaining megaspore resides in a tissue produced by cells of the megasporangium, the nucellus, the whole being surrounded by an integument. The integument is not complete, but, rather, leaves a passageway, the micropyle, through which the male gamete can gain access to the egg. The position of the micropyle is toward the central axis of the cone.
Some cells of the nucellus produce an exudate. This exudate, called a pollination drop, flows out to the mouth of the micropyle, where pollen grains adhere to the exudate. The pollination drop then shrinks and draws the pollen grains in through the micropyle to bring them into contact with the nucellus. The pollen is thus positioned an extremely short distance from where the egg will reside.
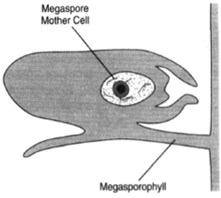 |
| Figure 27-3 A megasporangium, showing the megaspore mother cell before meiosis. The megasporangium is borne upon the upper surface of the megasporophyll. |
Some cells of the nucellus produce an exudate. This exudate, called a pollination drop, flows out to the mouth of the micropyle, where pollen grains adhere to the exudate. The pollination drop then shrinks and draws the pollen grains in through the micropyle to bring them into contact with the nucellus. The pollen is thus positioned an extremely short distance from where the egg will reside.
At the time of pollination, the female gametophyte has not yet grown to maturity; yet the pollen tube begins its slow growth through the nucellus. Digestive enzymes are secreted to create a channel through the nucellar tissue. The megaspore has produced the female gametophyte by numerous divisions. Toward the micropylar end of the gametophyte are two or more archegonia, each of which bears an egg cell. The growth of the pollen tube is very slow, taking as long as one year to reach the egg, which matures during this passage of time. The generative cell divides to form a stalk cell and a body cell. The body cell then divides to produce two nonmotile sperm nuclei. When the pollen tube reaches the archegonium, it ruptures and releases the sperm nuclei, one of which can fertilize the egg.
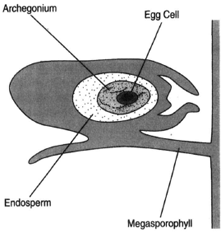 |
| Figure 27-4 The fully developed female gametophyte. |
The zygote undergoes mitoses without initially forming cell membranes. At the eight-cell stage, cell membranes form around most of the nuclei. At the sixteen-cell stage, the cells are arranged in tiers. The cells in the second tier elongate. These are the suspensor cells, and their elongation pushes the lowest group of four cells down through the base of the archegonium and into a nutritive tissue called the endosperm. These four cells represent embryos; thus, there are four one-celled embryos at this moment. Although all of them may begin growth, one will gain over the others, which will be unable to continue development
The growth of the embryo continues slowly, and the seeds may not be shed until after two or three years of development. Once shed, they may remain dormant for an extended period. The pine embryo is composed of a central hypocotyl, a radicle, a plumule, and several cotyledons.
Pines are described as evergreen, meaning that the leaves are not shed all at one time. Leaves remain on the tree varying lengths of time, depending on the species. Whereas Pinus strobus, the white pine, holds its leaves for two years, for example, Pinus aristata retains its leaves for as many as fourteen years. The leaves of P. aristata are long and slender (needle-like) and produced in bundles. The number of leaves in a bundle is usually three to five. Each leaf has a thick, waxy cuticle and sunken stomates, characteristics valuable in reducing water loss. Because leaves remain on the tree during winter, when water uptake is much reduced, the cuticle and sunken stomates thus represent important adaptations. A cross-sectional view of a pine leaf (figure 27-6) reveals a hypodermis two or three cells in thickness and immediately beneatht he epidermis. The hypodermis also helps prevent water loss. A vascular bundle or two is located at the center of the leaf. The bundle is surrounded by a parenchyma, which is boundebdy the endodermis. Outside the endodermis is a zone of mesophyll, through which resin ducts may be found passing.
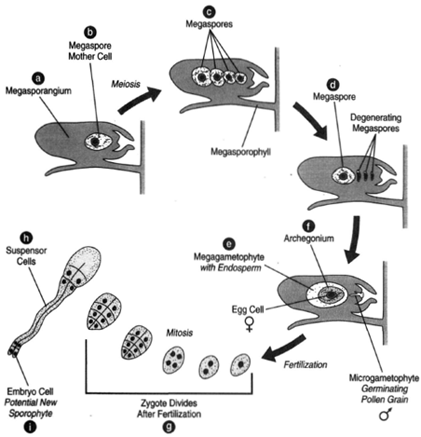 |
| Figure 27-5 Female gametophyte development and early embryo formation. (a) Megasporangium. (b) Megaspore mother cell. (c) Four megaspores are produce by meiosis. (d) Three of the megaspores degenerate, leaving only one. The megaspore grows into the female gametophyte having endosperm, (e), and archegonia, (f) each with an egg cell. (g) After fertilization, the zygote undergoes nuclear divisions. Here is shown the sixteen cell stage. (h) The second tier of cells, the suspensor cells, elongate. (i) Below the suspensor cells are embryo cells. Each embryo cell is a potential new sporophyte. |
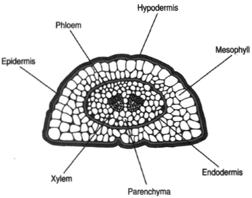 |
| Figure 27-6 Cross section of pine leaf. |
Before discussing the tissues of a pine stem, two terms need to be introduced: primary growth and secondary growth. Primary growth derives from cells at the tip of a stem, whereas secondary growth derives from a ring of cells a short distance below the tip (the cambium). A cross section of a stem near the apex, where no secondary growth has yet occurred, reveals a centrally located pith (parenchyma tissue) immediately outside of which is a circle of vascular strands and then a cortex bounded by an epidermis. A cross section of stem slightly below this reveals secondary tissues. A layer of cells between the primary xylem and primary phloem of the vascular strands as well as cells between the vascular strands collectively constitute the cambium, or actively dividing cells. Cambial cell divisions produthcee secondary vascular tissues, xylem and phloem. The xylem is deposited centrally to the cambium; the phloem, peripherally. The secondary xylem is composed of two kinds of cells: tracheids and fibers. In cross-sectional view, the tracheids formed during spring are relatively large compared to the smaller tracheids formed during autumn. This results in a clear delineation of annual rings in the wood. Numbers of resin ducts can be seen scattered throughout the wood, or secondary xylem.
As the secondary xylem increases, the cambium, which lies outside of this xylem, must move outward and increase its circumference. Cambium must therefore produce not only secondary xylem and secondary phloem, but also additional cambium cells. As the diameter of the stem increases by cambial activity, the cortex and epidermis (the outermost primary tissues) split and slough away.
A longitudinal section reveals rows of bordered pits in the walls of the tracheids. As indicated previously, tracheid cells have both primary and secondary cell walls. Whereas the pits are created by interruptions in the secondary wall, the primary wall is continuous through the pits. The primary wall in the region of the pit has a thickening, called the torus. The secondary wall overhangs the pit, thus making a bordered pit. The pits of adjacent cells are aligned, making a pit pair. The pit membrane is flexible and can be pressed one way or the other.
Support our developers




