Rusts
Puccinia graminis fungus has taken a large toll on wheat yields throughout history. An epidemic in 1916 caused the loss of 14 million bushels in the United States and 100 million bushels in Canada. In 1935, 60 percent of the crop was lost in Minnesota and the Dakotas; in other areas, the loss was near total. In 1953 to 1954, the loss was more than 80 percent in the same area. In Roman times, it was thought that damaged wheat was a visitation upon wicked persons. For hundreds of years it was known or suspected that the common barberry plant was in some way involved; yet not until 1800 did it become known that wheat rust is caused by a fungus. German mycologist Heinrich DeBary (1831-1888) is credited with working out the details of 'P. graminis' life cycle in 1865. The details are complicated, with five different kinds of spores being produced in the complete cycle (see figure 20-5). |
| Figure 20-3 Phallus impudicus, another stinkhorn. |
 |
| Figure 20-2 Dictyophora, a stinkhorn. Its odor makes it attractive to flies, which perch on the top. The fungus was fonerly used to make a salve for the treatment of rheumatism. (Illustration by Laurette Richin) |
The infection of the wheat plant in spring shows a rust-red zone, which, on examination with the microscope, reveals a cluster of sporangia. The cluster is called a sorus, and the spores that are produced here have two nuclei. These are called uredospores. Air currents carry the uredospores, thus spreading the infection to other wheat plants. In late summer, the nuclei fuse to create a diploid cell. These nuclei then divide, and a cross wall forms, thereby creating two cells at the end of a stalk. In this stage, the sorus becomes black, and the spores are called teliospores. Teliospores are thick walled and generally do not germinate until the following spring. They remain on the straw and may germinate in the field or, perhaps, the manure pile. Germination produces an elongated basidium partitioned by walls. The process involves meiosis yielding four haploid basidiospores . As shown in figure 20-5, the basidiospores are suspended on the ends of pointed projections called the sterigmata (sterigma). There are two types of basidiospores: plus (+) and minus (−).
There is only one place that a basidiospore can germinate: on the surface of a leaf of the common barberry, Berberis vulgaris. The infection is highly host specific; there is no other place that it can grow. It is so specific, in fact, that it is able to grow only on the upper surface of the leaf. A mycelium is produced in the tissue of the leaf. After a time, pockets are formed on the upper surface. These pockets are called spermogonia (or pycnia), and spermatia are produced in them. Two kinds of spermatia are produced from the
 |
| Figure 20-4 End wall perforation |
plus and minus teliospores: plus and minus. The spermogonia secrete a sugary fluid attractive to insects, which by seeking this nectar, bring mating types of spermatia together. Although plus and minus spermatia are thereby paired, nuclear fusion does not occur. The cells fuse, but the nuclei do not; a dkaryotic condition thus results. The dikaryotic cells then produce cup-shaped aecia (aecium) on the underside of the barberry leaves. Binucleated aeciosporeasr e then produced in these aecia chains. These aeciospores arteh e agents able to infect the wheat plant, and they mark the end of the life cycle.
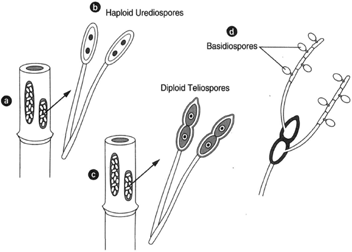 |
| Figure 20-5 Puccinia graminis (wheat rust) infection on the wheat plant. At left, (a) redstage sorus occurring in spring. (b) The binucleated spores are haploid uredospores. At right of the red-stage sorus, the black-stage sorus (c) occurring in the fall. Diploid, 2N teliospores are formed. At far right, a teliospore germinates to produce basidiospores, (d). |
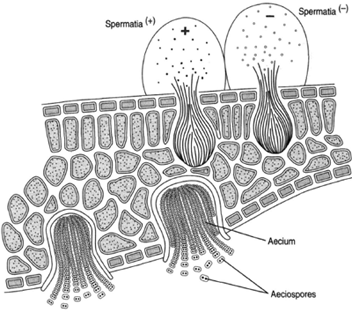 |
| Figure 20-6 Puccinia graminis infection on barberry leaf. On the upper surface of the leaf, spermogonia with spermatia, which grow from teliospores. Uninucleated spermatia become paired without nuclear fusion, producing aecia on the lower surface of the leaf. The aeciospores are binucleated. |
Recall now that the early-season infection of the wheat produces binucleated uredospores, and that nuclear fusion is long delayed. The nuclei do not fuse until late summer, whtehne black sorus stage occurs and teliospores are produced. Note also that there is an alternation of generations, an alternation of haploid cells and diploid cells, and an alternation of hosts, called heteroecism.
Although some species of rust fungi are able to complete the life cycles by parasitizing a single host, large numbers of rusts require two hosts. The white pine blister rust Cronartium ribicola, responsible for the loss of millions of trees, infects white pine, with the alternate host being gooseberries or currents. Some measure of control has been attained by eradicating gooseberry and current plants in regions where white pine is an important lumber. In the same way, barberry eradication has met with modest success in controlling wheat rust.
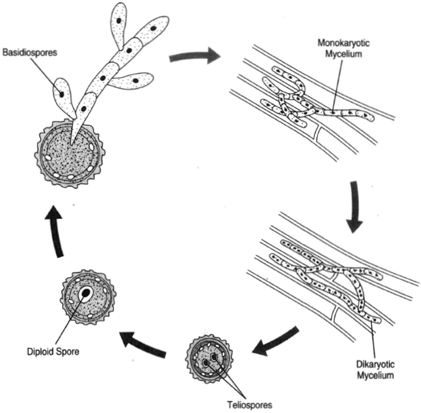 |
| Figure 20-7 Ustilago maydis, corn smut. Basidiospores infect the cells of the corn plant, producing a monokaryotic mycelium, which later becomes dikaryotic. The dikaryotic cells produce teliospores. The nuclei fuse, producing a diploid spore, which produces a basidium by meiosis. |
Support our developers




