Concentrations of Calcium in Plants
Forms of Calcium Compounds
Within plants, calcium is present as Ca2+ ions attached to carboxyl groups on cell walls by cation-exchange reactions. As approximately one third of the macromolecules in the primary cell wall are pectin (9), it can be seen that a large proportion occurs as calcium pectate. Pectin may also join with anions, such as vanadate, and serve to detoxify these ions. The Ca2+ cation will also join with the organic anions formed during the assimilation of nitrate in leaves; these anions carry the negative charge that is released as nitrate is converted into ammonium (46). Thus, there will be formation of calcium malate and calcium oxalacetate and, also very commonly, calcium oxalate in cells.Calcium oxalate can occur within cells and as extracellular deposits. In a study of 46 conifer species, all contained calcium oxalate crystals (47). All of the species in the Pinaceae family accumulated the compound in crystalliferous parenchyma cells, but the species not in the Pinaceae family had the compound present in extracellular crystals.
This accumulation of calcium oxalate is common in plants in most families. Up to 90% of total calcium in individual plants is in this form (48,49). Formation of calcium oxalate crystals occurs in specialized cells, crystal idioblasts, and as the calcium oxalate in these cells is osmotically inactive their formation serves to lower the concentration of calcium in the apoplast of surrounding cells without affecting the osmotic balance of the tissue (48). A variety of different forms of the crystals occur (49), and they can be composed of calcium oxalate monohydrate or calcium oxalate dihydrate (50).
Distribution of Calcium in Plants
Calcium moves toward roots by diffusion and mass flow (51,52) in the soil. A number of calciumspecific ion channels occur in the membranes of root cells, through which influx occurs, but these channels appear to be more involved in enabling rapid fluxes of calcium into the cytoplasm and organelles as part of signalling mechanisms (53). This calcium is then moved into vacuoles, endoplasmic reticulum, or other organelles, with movement occurring by means of calcium-specific transporters (20).The bulk entry of calcium into roots occurs initially into the cell walls and in the intercellular spaces of the roots, giving a continuum between calcium in the soil and calcium in the root (54). For calcium to move from the roots to the rest of the plant, it has to enter the xylem, but the Casparian band of the endodermis is an effective barrier to its movement into the xylem apoplastically. However, when endodermis is first formed, the Casparian band is a cellulosic strip that passes round the radial cell wall (state I endodermis), so calcium is able to pass into the xylem if it passes into the endodermal cells from the cortex and then out again into the pericycle, through the plasmalemma abutting the wall (55). This transport seems to occur, with the calcium moving into the endodermal cells (and hence into the symplasm) through ion channels and from the endodermis into the pericycle (and ultimately into the much higher concentration of calcium already present in the xylem) by transporters (56,57). Highly developed endodermis has suberin lamellae laid down inside the cell wall around the entire cell (state II endodermis), and in the oldest parts of the root, there is a further layer of cellulose inside this (state III) (55). Although some ions such as K+ can pass through state II endodermal cells, Ca2+ cannot. There are plasmodesmata between endodermis and pericycle cells, even where the Casparian band is well developed, but although phosphate and K+ ions can pass, the plasmodesmata are impermeable to Ca2+ ions.
This restriction in effect limits the movement of calcium into the stele to the youngest part of the root, where the endodermis is in state I. Some movement occurs into the xylem in older parts of the root, and this transport can occur by two means. It is suggested that movement of calcium through state III endodermis might occur where it is penetrated by developing lateral roots, but the Casparian band rapidly develops here to form a complete network around the endodermal cells of the main and lateral roots (55). The second site of movement of calcium into the stele is through passage cells (55). During the development of state II and state III endodermis some cells remain in state I. These are passage cells. They tend to be adjacent to the poles of protoxylem in the stele, and they are the site of calcium movement from cortex to pericycle.
In some herbaceous plants (e.g., wheat, barley, oats), the epidermis and cortex are lost from the roots, especially in drought, so the passage cells are the only position where the symplast is in contact with the rhizosphere (55). Most angiosperms form an exodermis immediately inside the epidermis, and the cells of this tissue also develop Casparian bands and suberin lamellae, with passage cells in some places (55). These passage cells are similarly the only place where the symplasm comes in contact with the rhizosphere.
Because of this restricted entry into roots, calcium enters mainly just behind the tips, and it is mostly here that it is loaded into the xylem (Figure 5.6). Absorption of calcium into the roots may be passive and dependent on root cation-exchange capacity (CEC) (58). Transfer of calcium into roots is hardly affected by respiratory uncouplers, although its transfer into the xylem is affected (54,59).
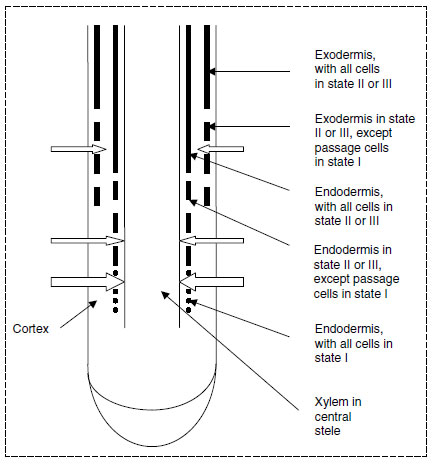 |
| FIGURE 5.6 Diagrammatic representation of longitudinal section of root, showing development of endodermis and exodermis, and points of entry of calcium. (Based on C.A. Peterson and D.E. Enstone, Physiol. Plant 97: 592-598, 1996.) |
Once in the xylem the calcium moves in the transpiration stream, and movement around the plant is restricted almost entirely to the xylem (60,61) as it is present in the phloem only at similarly low concentrations to those that occur in the cytoplasm.
As calcium is not mobile in the phloem, it cannot be retranslocated from old shoot tissues to young tissues, and its xylem transport into organs that do not have a high transpiration rate (such as fruits) is low (22). Its flux into leaves also declines after maturity, even though the rate of transpiration by the leaf remains constant (62), and this response could be related to a decline in nitrate reductase activity as new leaves in the plant take over a more significant assimilatory role (22,63). When a general deficiency of calcium occurs in plants, because of the low mobility of calcium in phloem, it is the new leaves that are affected, not the old leaves, as calcium in a plant remains predominantly in the old tissues (Figure 5.7).
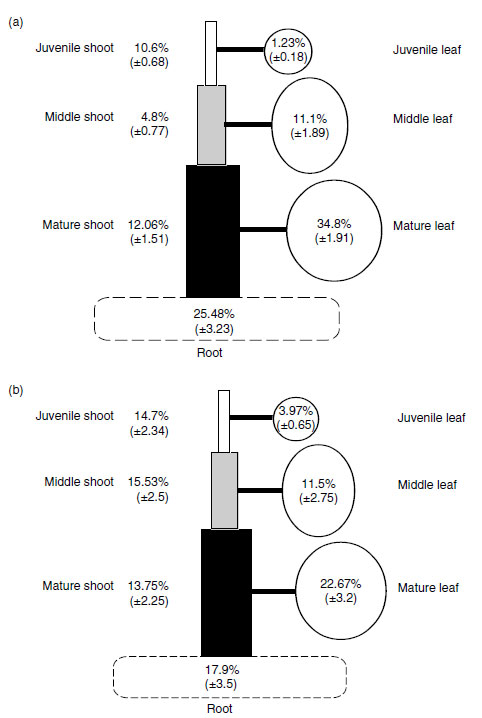 |
| FIGURE 5.7 Distribution of calcium (a) and distribution of dry mass (b) in Capsicum annuum cv Bendigo plants grown for 63 days in nutrient solution (values are means of values for nine plants ± standard error). |
It was long thought that a direct connection occurs between the amount of transpiration that a plant carries out and the amount of Ca2+ that it accumulates. For example, in a study of five tomato cultivars grown at two levels of electrical conductivity (EC) there was a linear, positive relationship between water uptake and calcium accumulation over 83 days (64). However, with the movement of Ca2+ in the symplasm of the endodermis apparently being required for xylem loading, it became accepted that Ca2+ is taken up in direct proportion to plant growth, as new cation-exchange sites are made available in new tissue. The link with transpiration could therefore be incidental, because bigger plants transpire more. Thus the plant acts as a giant cation exchanger, taking up calcium in proportion to its rate of growth. Supplying calcium to decapitated plants at increased ion activity (concentration) leads to increased uptake of the ion, a process that appears to contradict this concept. However, in intact plants, the rate of uptake is independent of external ion activity, as long as the ratios of activities of other cations are constant relative to the activity of Ca2+ (65,66).
The theory that calcium travels across the root in the apoplastic pathway, until it reaches the Casparian band of the endodermis and at which its passage to the xylem becomes symplastic, is not entirely without problems. White (56,67) calculated that for sufficient calcium loading into xylem, there must be two calcium-specific ion channels per �m2 of plasmalemma on the cortex side of the endodermis. This possibility is plausible. However, for the flux of calcium to continue from the endodermis into the pericycle there must be 0.8 ng Ca2+-ATPase protein per cell, equivalent to 1.3 mg per gram of root fresh weight. This concentration is greater than the average total root plasmalemma protein concentration in plants. Furthermore, there is no competition between Ca2+, Ba2+, and Sr2+ for transport to mouse-ear cress (Arabidopsis thaliana Heynh.) shoots, as would be expected if there was protein-mediated transport in the symplast. Some apoplastic transport to the xylem cannot be ruled out.
The walls of xylem vessels have cation-exchange sites on them; in addition to the whole plant having a CEC, the xylem represents a long cation-exchange column with the Ca2+ ions moving along in a series of jumps (54). The distance between each site where cation exchange occurs depends on the velocity of the xylem sap and the concentration of Ca2+ ions in it (54). Thus, for transpiring organs such as mature leaves, the calcium moves into them quickly, but for growing tissues such as the areas close to meristems, the supply of calcium is dependent on the deposition of cell walls and the formation of new cation-exchange sites (54). It has been suggested that transpiring organs receive their calcium in the transpiration stream during the day, and growing tissues receive their calcium as a result of root pressure during the night (54).
The restriction in movement of calcium to the xylem gives rise to most of the calcium-deficiency disorders in plants. For example, BER (Figure 5.4 and Figure 5.5) in tomatoes occurs because the developing fruits are supplied solutes better by phloem than by xylem as the fruits do not transpire. Xylem fluid goes preferentially to actively transpiring leaves, giving a lower input of calcium into developing fruits (68). A period of hot, sunny weather not only gives rise to so much transpiration that calcium is actively pulled into leaves, but gives rates of photosynthesis that are enhanced to the extent that fruits expand very rapidly. Under these conditions, it is likely that localized deficiencies of calcium will occur in the distal end of the fruits, furthest from where the xylem enters them (the 'blossom' end) (Figure 5.4 and Figure 5.5). Typically, tomatoes grown for harvest in trusses are more susceptible to BER than 'single-pick' types, presumably because the calcium has to be distributed to several developing sinks at the same time. Conditions that promote leaf transpiration, such as low humidity, lower the import of calcium into developing fruits and increase the risk of BER.
It has also been thought in the past that salinity, which increases water potential in the root medium, would likewise restrict calcium import into the fruit, accounting for increased incidence of BER that is known to occur under saline conditions. This effect of salinity could be important in some natural soils, but is also important in glasshouse production of tomatoes as high-electroconductivity (EC) nutrient solutions are sometimes used because they increase dry matter production in fruits and improve flavor. However, it has been observed that if the ion activity ratios aK/√(aCa + aMg) and aMg/aCa are kept below critical values, the risks of BER developing in high-EC nutrient solutions are lowered (69). It seems as if one of the causes of increased BER with salinity is normally due to increased uptake of K+ and Mg2+, which restricts the uptake and distribution of Ca2+ ions.
Cultivars differ in susceptibility to BER, with beefsteak and plum types of tomato being particularly susceptible. Susceptibility is related partly to fruit yield, and two susceptible cultivars of tomato (Calypso and Spectra) were shown to have a higher rate of fruit set than a nonsusceptible cultivar (Counter) (70). The so-called calcium-efficient strains of tomato do not have lower incidence of BER, since although they accumulate more dry matter than Ca-inefficient strains, this accumulation is predominantly in the leaves (64). Cultivars with relatively small fruits, such as Counter (70), and with xylem development in the fruit that is still strong under saline conditions (71), are able to accumulate comparatively high proportions of their calcium in the distal end of the fruits under such conditions and are less susceptible to BER (64). However, cultivars with low yields of fruits per plant may show even lower incidence of BER than those with high yields (64).
Losses of tomatoes to BER in commercial horticulture can reach 5% in some crops, representing a substantial loss of potential income. The main approaches to prevent BER are to use less-susceptible cultivars and to cover the south-facing side of the glasshouse (in the northern hemisphere) with white plastic or whitewash to limit the amount of solar radiation of the nearest plants and prevent their fruits from developing too quickly in relation to their abilities to accumulate calcium.
Calcicole and Calcifuge Species
In general, calcicole species contain high concentrations of intracellular calcium, and calcifuge species contain low concentrations of intracellular calcium. The different geographic distributions of these plants seem to be largely determined by a range of soil conditions other than just calcium concentration in the soil per se. In the calcareous soils favored by calcicoles, in addition to high concentration of Ca2+, pH is high, giving low solubility of heavy metal ions and high concentrations of nutrient and bicarbonate ions. In contrast, the acid soils favored by calcifuges have low pH, high solubility of heavy metal ions, and low availability of nutrients (5).The growth of calcicole species is related strongly to the concentration of calcium in the soil, but the inability of calcicole species to grow in acid soils is linked strongly to an inability to tolerate the high concentrations of ions of heavy metals, in particular Al3+, Mn2+, and Fe3+ (5,72). For calcifuge species, the difficulty in growing in a calcareous soil stems from an inability to absorb iron, although in some calcareous soils low availability of phosphate may also be a critical factor. In an experiment with tropical soils in which the sorption of phosphate from Ca(H2PO4)2 solution (and its subsequent desorption) were measured, pretreating the soil with calcium sulfate solution increased the sorption of phosphate (73). In the most acid of the soils tested, sorption of phosphate was increased by 93%. Because the extracts of the soil became more acid following calcium sulfate treatment, it appears that the calcium was attracted to the sites previously occupied by H+ ions, and when present, itself offered more sites for sorption of phosphate ions. Where the supply of phosphorus to plants is limited because it is sorbed to soil inorganic fractions, it seems as if sorption to calcium is more difficult to break than sorption to other components. In an experiment in which wheat (Triticum aestivum L.) and sugar beet (Beta vulgaris L.) were grown in a fossil Oxisol, with mainly Fe/Al-bound P, and in a Luvisol, a subsoil from loess with free CaCO3 and mainly Ca-bound P, both species (but particularly the sugar beet) were able to mobilize the Fe/Albound P more than the Ca-bound P (74).
Some plants are much more efficient than others at taking up phosphate from calcium-bound pools in the soil. One efficient species is buckwheat (Fagopyrum esculentum Moench). In a comparison of this species and wheat, the buckwheat took up 20.1 mg P per pot compared with 2.1 mg P per pot for wheat if nitrogen was supplied as nitrate (75). Changing the nitrogen supply to ammonium nitrate increased phosphorus accumulation by the wheat largely, with very little effect on the buckwheat, indicating that it is the capacity of buckwheat to acidify the rhizosphere even when the nitrogen supply is nitrate that makes buckwheat able to utilize this firmly bound source of phosphorus.
For calcifuge species growing on calcareous soils, it seems as if the availability of iron is the most significant factor affecting plant growth, with chlorosis occurring due to iron deficiency. However, this deficiency is caused largely by immobilization of iron within the leaves, not necessarily a restricted absorption of iron (76,77). Calcicole species seem to make iron and phosphate available in calcareous soils by exudation of oxalic and citric acids from their roots (78). The high concentrations of bicarbonate ions in calcareous soils seem to be important in inhibition of root elongation of some calcifuge species (79).
Critical Concentrations of Calcium
The concentrations of calcium in plants are similar to the concentrations of potassium, in the range 1 to 50 mg Ca g-1 dry matter (Mengel, this volume). Most of the calcium is located in the apoplast, and where it is present in the symplast, it tends to be stored in organelles or vacuoles or is bound to proteins. The concentration of free Ca2+ in a root cortical cell is of the order of 0.1 to 1.0 mmol m-3 (54).In general, monocotyledons contain much less calcium than dicotyledons. In an experiment comparing the growth of ryegrass (Lolium perenne L.) and tomato, the ryegrass reached its maximum growth rate when the concentration of calcium supplied gave a tissue concentration of 0.7mg g-1 dry mass, whereas tomato reached its maximum growth rate only when tissue concentration was 12.9 mg g-1 (80,81). This difference between monocotyledons and dicotyledons is dictated by the CEC of the two groups of plants. In algal species, where the cell wall is absent and CEC is consequently low, calcium is required only as a micronutrient (82).
Tissue concentrations of calcium can vary considerably according to the rate of calcium supply. In a study by Loneragan and Snowball (81), internal Ca2+ concentrations were reasonably constant for 0.3, 0.8, and 2.5 �M calcium in the flowing nutrient solutions for each plant species tested, but with 10, 100, or 1000 �M Ca2+ supply, internal Ca2+ concentrations were noticeably higher. In a recent study of chickpea (Cicer arietinum L.), nine different Kabuli (large-seeded) accessions had a mean concentration of Ca2+ in nodes 4 to 7 of the shoot of 17.4 mg g-1 dry mass after 33 days of growth, and 10 different Desi (small-seeded) accessions had a mean Ca2+ concentration of 17.1 mg g-1 dry mass (83). In the Kabuli accessions, the range was between 13.5 and 20.6 mg g-1, compared with between 13.1 and 19.0 mg g-1 in the Desi accessions, so different genotypes of the same species grown under the same conditions seem to contain very similar shoot calcium concentrations.
There are considerable amounts of data regarding what the critical concentrations of calcium are in different plants and different species. For data on these concentrations in a large number of species, the reader is referred to some special publications (84,85).
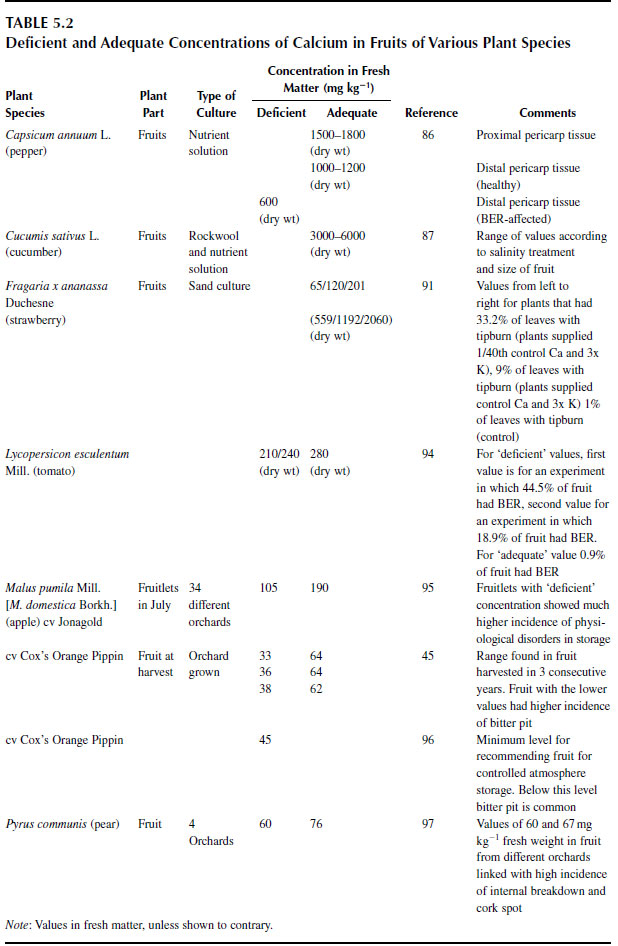
In a study of three cultivars of bell pepper, mean tissue concentrations ranged only from 1.5 to 1.8 mg g-1 dry mass in the proximal parts and from 0.95 to 1.3 mg g-1 dry mass in the distal part of healthy fruits. concentrations in fruits suffering BER were between 0.6 and 1.0 mg g-1 (86). Concentrations of calcium in fruits of cucumber (Cucumis sativus L.), a plant that is not susceptible to BER, are typically three to seven times these values (87).
There is one important exception to the finding that internal calcium concentrations are relatively constant regardless of how plants are grown. Plants supplied with nitrogen as ammonium tend to have much lower concentrations of cations, including calcium, than plants supplied with nitrate (22). Thus, tomato plants supplied with ammonium-N are more prone to BER than plants grown on nitrate.
Tabulated Data of Concentrations by Crops
Concentrations of Ca2+ in shoots and fruits of some crop species are reported in Table 5.1 and Table 5.2.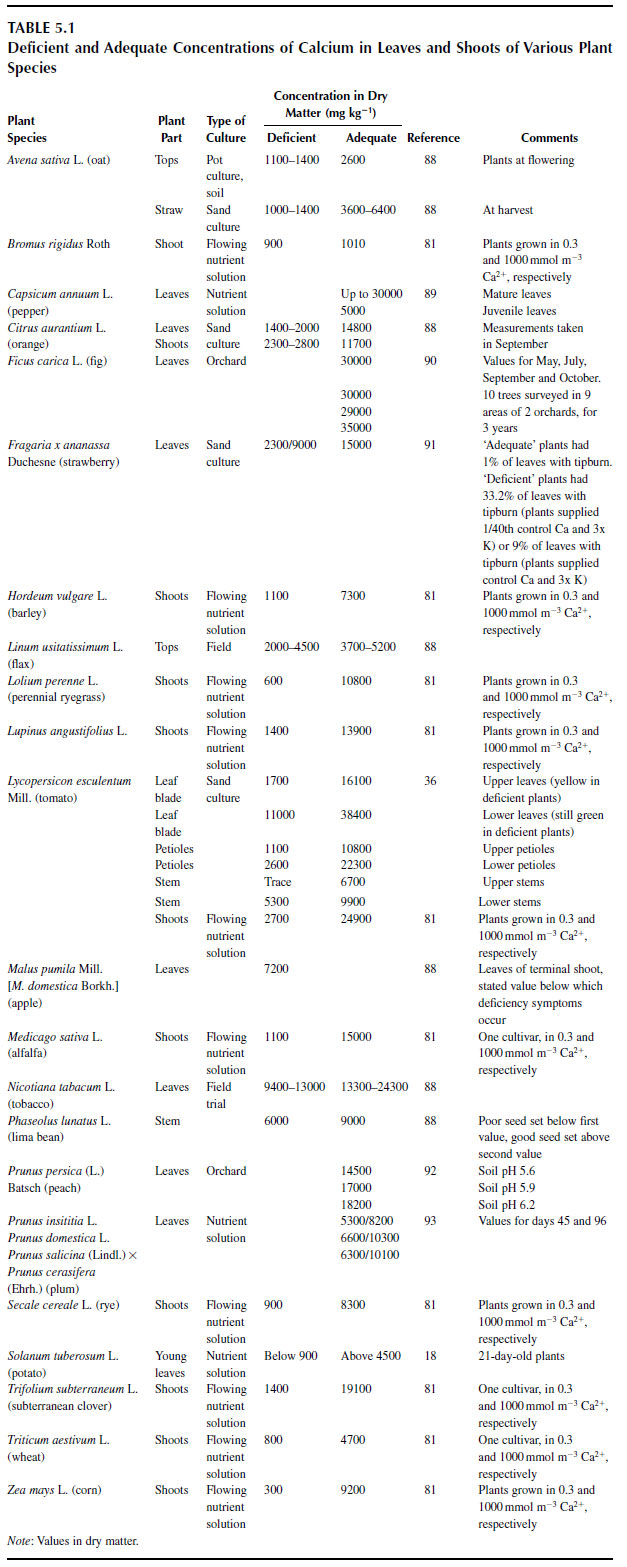 |
 |
Support our developers




