Pollen
Contents
» The structure of pollen» Pollination
» Pollen as a carrier of ecological information in plants
» Pollen in the fossil record
» Hay fever
» Pollen in human diets
» References
The structure of pollen
Each pollen grain contains vegetative (non-reproductive) cells (only a single cell in most flowering plants but several in other seed plants) and a generative (reproductive) cell containing two nuclei: a tube nucleus (that produces the pollen tube) and a generative nucleus (that divides to form the two sperm cells). The group of cells is surrounded by a cellulose-rich cell wall called the intine, and a resistant outer wall composed largely of sporopollenin called the exine.Pollen is produced in the microsporangium (contained in the anther of an angiosperm flower, male cone of a coniferous plant, or male cone of other seed plants). Pollen grains come in a wide variety of shapes, sizes, and surface markings characteristic of the species (see Electron micrograph at top right). Most, but certainly not all, are spherical. Pollen grains of pines, firs, and spruces are winged. The smallest pollen grain, that of the Forget-me-not (Myosotis spp.), is around 6 µm (0.006 mm) in diameter. Wind-borne pollen grains can be as large as about 90-100 µm. The study of pollen is called palynology and is highly useful in paleoecology, paleontology, archeology, and forensics.
Pollen grains may have furrows, the orientation of which (relative to the original tetrad of microspores) classify the pollen as colpate or sulcate. The number of furrows or pores helps classify the flowering plants, with eudicots having three colpi (tricolpate), and other groups having one sulcus.
Except in the case of some submerged aquatic plants, the mature pollen-grain has a double wall, a thin delicate wall of unaltered cellulose (the endospore or intine) and a tough outer cuticularized exospore or exine. The exine often bears spines or warts, or is variously sculptured, and the character of the markings is often of value for identifying genus, species, or even cultivar or individual. In some flowering plants, germination of the pollen grain often begins before it leaves the microsporangium, with the generative cell forming the two sperm cells.
Pollination
The transfer of pollen grains to the female reproductive structure (pistil in angiosperms) is called pollination. This transfer can be mediated by the wind, in which case the plant is described as anemophilous (literally wind-loving). Anemophilous plants typically produce great quantities of very lightweight pollen grains, sometimes with air-sacs. Non-flowering seed plants (e.g. pine trees) are characteristically anemophilous.Anemophilous flowering plants generally have inconspicuous flowers. Entomophilous (literally insect-loving) plants produce pollen that is relatively heavy, sticky and protein-rich, for dispersal by insect pollinators attracted to their flowers. Many insects and some mites are specialized to feed on pollen, and are called palynivores.
In non-flowering seed plants, pollen germinates in the pollen chamber, located beneath and inside the micropyle. A pollen tube is produced, which grows into the nucellus to provide nutrients for the developing sperm cells. Sperm cells of Pinophyta and Gnetophyta are without flagella, and are carried by the pollen tube, while those of Cycadophyta and Ginkgophyta have many flagella.
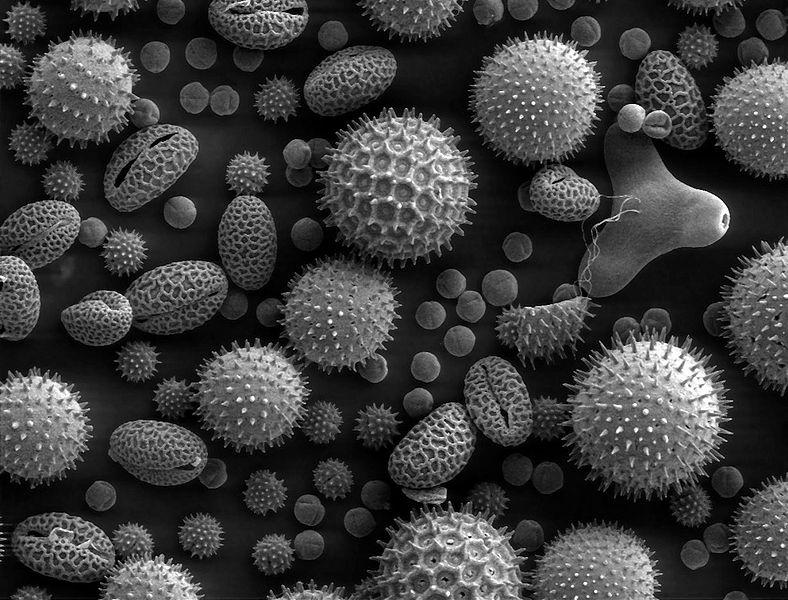
SEM image of pollen grains from a variety of common plants: sunflower (Helianthus annuus), morning glory (Ipomoea purpurea), prairie hollyhock (Sidalcea malviflora), oriental lily (Lilium auratum), evening primrose (Oenothera fruticosa), and castor bean (Ricinus communis).

Closeup image of a cactus flower and its stamens

Honey bee on sedum with pollen basket

Pollen from Phacelia (in purple)

Pollen sticking to a bee. Insects involuntarily transporting pollen from flower to flower play an important role in many plants' reproductive cycles.

Marmelade fly sitting on a grey-haired rockrose, its face and legs covered in pollen.

Clumps of yellow pollen on a flower head.
Pollen as a carrier of ecological information in plants
A Russian theoretical biologist, Vigen Geodakyan (Geodakian), has suggested that the quantity of pollen reaching a pistillate flower can transmit ecological information and also regulate evolutionary plasticity in cross-pollinating plants. Plentiful pollen indicates optimum environmental conditions (for example a plant that is situated at the center of its natural range, in ideal growing conditions, with a large number of male plants nearby, and favorable weather conditions), whereas a small amount of pollen indicates extreme conditions (at the borders of its range, with a deficiency of male plants, and adverse weather conditions). Geodakian believes that the quantity of pollen reaching a pistillate flower defines the sex ratio, dispersion and sexual dimorphism of a plant population. High pollen quantity leads to a reduction of these characteristics and stabilization of a population. Small quantity leads to their increase and destabilization of a population.Dependence of the secondary sex ratio on the amount of fertilizing pollen was confirmed on four dioecious plant species from three families — Rumex acetosa (Polygonaceae), Melandrium album (Cariophyllaceae), Cannabis sativa and Humulus japonicus (Cannabinaceae).
Dependence of offspring phenotype variety on amount of pollen was observed by Ter-Avanesyan in 1949. All three studied species of plants (cotton plant, black-eyed pea, and wheat) showed dependence in the direction forecast by the theory — fertilization with a small amount of pollen resulted in an increase in the diversity of the offspring. Ter-Avanesian writes that as a result of a limited pollination “instead of homogenous sorts we get populations”.
Pollen in the fossil record
Pollen's sporopollenin outer sheath affords it some resistance to the rigours of the fossilisation process that destroy weaker objects; it is also produced in huge quantities. As such, there is an extensive fossil record of pollen grains, often disassociated from their parent plant. The discipline of palynology is devoted to the study of pollen, which can be used both for biostratigraphy and to gain information about the abundance and variety of plants alive — which can itself yield important information about paleoclimates. Pollen is first found in the fossil record in the late Devonian period and increases in abundance until the present day.Hay fever
Allergy to pollen is called hay fever. Generally pollens that cause allergies are those of anemophilous plants (pollen is dispersed by air currents.) Such plants produce large quantities of lightweight pollen (because wind dispersal is random and the likelihood of one pollen grain landing on another flower is small) which can be carried for great distances and are easily inhaled, bringing it into contact with the sensitive nasal passages.In the US, people often mistakenly blame the conspicuous goldenrod flower for allergies. Since this plant is entomophilous (its pollen is dispersed by animals), its heavy, sticky pollen does not become independently airborne. Most late summer and fall pollen allergies are probably caused by ragweed, a widespread anemophilous plant.
Arizona was once regarded as a haven for people with pollen allergies, although several ragweed species grow in the desert. However, as suburbs grew and people began establishing irrigated lawns and gardens, more irritating species of ragweed gained a foothold and Arizona lost its claim of freedom from hay fever.
Anemophilous spring blooming plants such as oak, birch, hickory, pecan, and early summer grasses may also induce pollen allergies. Most cultivated plants with showy flowers are entomophilous and do not cause pollen allergies.
Pollen in human diet
A variety of producers have started selling pollen for human consumption, often marketed as a food (rather than a dietary supplement). The largest constituent is carbohydrates, with protein content ranging from 7 to 35 percent depending on the plant species collected by bees.The U.S. Food and Drug Administration (FDA) has not found any harmful effects of pollen consumption, except from the usual allergies. However, FDA does not allow pollen marketers in the United States to make health claims about their produce, as no scientific basis for these has ever been proved. Furthermore, there are possible dangers not only from allergic reactions but also from contaminants such as pesticides and from fungi and bacteria growth related to poor storage procedures. A manufacturers's claim that pollen collecting helps the bee colonies is also controversial.
References
- Pleasants, Jm; Hellmich, Rl; Dively, Gp; Sears, Mk; Stanley-Horn, De; Mattila, Hr; Foster, Je; Clark, P; Jones, Gd (October 2001). "Corn pollen deposition on milkweeds in and near cornfields". Proceedings of the National Academy of Sciences of the United States of America 98 (21): 11919–24.- Kenneth R. Sporne (1972). "Some Observations on the Evolution of Pollen Types in Dicotyledons". New Phytologist 71 (1): 181–185.
- Walter S. Judd and Richard G. Olmstead (2004). "A survey of tricolpate (eudicot) phylogenetic relationships". American Journal of Botany 91: 1627–1644.
- Geodakyan V. A. (1977). The Amount of Pollen as a Regulator of Evolutionary Plasticity of Cross-Pollinating Plants. “Doklady Biological Sciences” 234 N 1-6, 193–196.
- Соrrеns С. (1922) Geschlechtsbestimmung und Zahlenverhaltnis der Geschlechter beim Sauerampfer (Rumex acetosa). “Biol. Zbl.” 42, 465-480.
- Rychlewski J., Kazlmierez Z. (1975) Sex ratio in seeds of Rumex acetosa L. as a result of sparse or abundant pollination. “Acta Biol. Cracov” Scr. Bot., 18, 101-114.
- Correns C. (1928) Bestimmung, Vererbung und Verteilung des Geschlechter bei den hoheren Pflanzen. Handb. Vererbungswiss., 2, 1-138.
- Riede W. (1925) Beitrage zum Geschlechts- und Anpassungs-problem. “Flora” 18/19
- Kihara H., Hirayoshi J. (1932) Die Geschlechtschromosomen von Humulus japonicus. Sieb. et. Zuce. In: 8th Congr. Jap. Ass. Adv. Sci., p. 363—367 (cit.: Plant Breeding Abstr., 1934, 5, № 3, p. 248, ref. № 768).
- Geodakyan, V.A. & Geodakyan, S.V., (1985) Is there a negative feedback in sex determination? “Zurnal obschej biol.” 46 201-216 (in Russian). ).
- Ter-Avanesyan D. V. (1949). Tr. Prikl. Bot, Genet, Selekt., 28 119.
- Ter-Avanesian D. V. (1978) Significance of pollen amount for fertilization. “Bull. Torrey Bot. Club.” 105 N 1, 2–8.
- Sanford, citing P. Witherell, "Other Products of the Hive," Chapter XVIII, The Hive and the Honey Bee, Dadant & Sons, Inc., Hamilton, IL, 1975
- Malcolm T. Sanford. "Producing Pollen". University of Florida, Institute of Food and Agricultural Sciences.
Support our developers




