Effect of chemical mutagens on nucleotide sequence
(a) Incorporation of base analogues:
5-bromouracil (5-Bu), 5-chlorouracil (5-Cu) and 5-iodouracil (5-Iu) can replace thymine in DNA but, 2-aminopurine (2-AP) is incorporated in such a small amount that it could not be possible to find out which base it replaces. 2,6-diaminopurine is also highly mutagenic, but its effect has not been studied in detail. Although these base analogues are much less inhibitory for bacterial and phage growth than other purine and pyrimidine analogues, yet they are much more mutagenic. Their mutagenic effect has been attributed, therefore, to pairing mistakes due to their incorporation in DNA.
(i) 5-Bromouracil (Bu) or 5-Bromodeoxy-uridine (BUdr). 5-Bu can pair with adenine just like thymine (Fig. 23.2). Occasionally 5-Bu loses a hydrogen atom in position-1 and pairs with guanine instead of adenine (Fig. 23.3). This pairing mistake may occur at the time of 'incorporation' (Fig. 23.4) or after incorporation at the time of 'replication'.
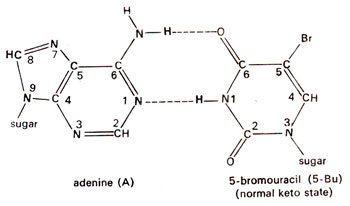
Fig. 23.2. Base pairing between adenine (A) and normal keto state of 5-bromouracil (5-Bu).
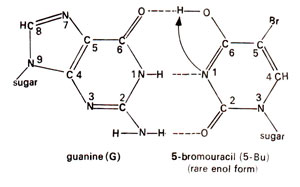
Fig. 23.3. Base pairing between adenine (A) and rare enol form of 5-bromouracil (5-Bu).
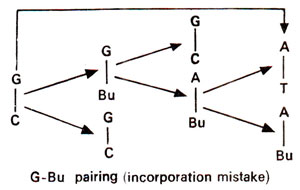
Fig. 23.4. Steps involved in GC →AT base pair transition due to mistake in incorporation of 5-bromouracil (5-Bu).
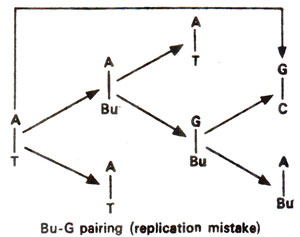
Fig. 23.5. Steps involved in AT → GC base pair transition due to mistake in replication caused by the presence of 5-bromouracil (5-Bu).
Bromodeoxyuridine (BUdR) is more effective mutagen probably because it is more easily converted to deoxyribonucleoside triphosphate. Both Bu and BUdR induce point mutations.
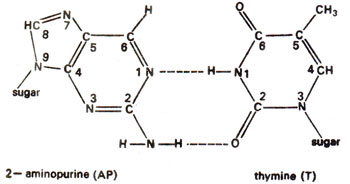
Fig. 23.6. Normal base pairing between 2-aminopurine (2-AP) and thymine (T).
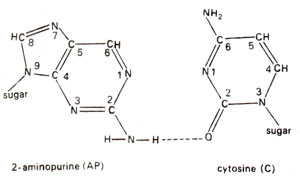
Fig. 23.7. Rare base pairing between 2-aminopurine (2-AP) and cytosine (C).
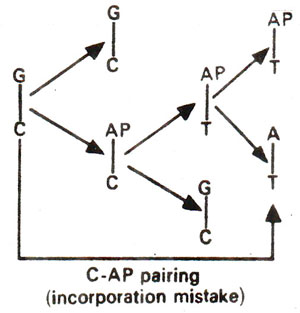
Fig. 23.8. Steps involved in GC →AT base pair transition due to mistake in incorporation of 2-aminopurine (AP).
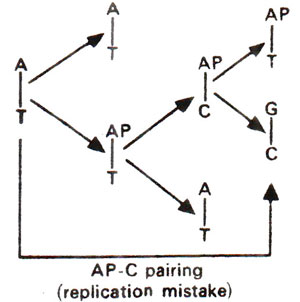
Fig. 23.9. Steps involved in AT →GC base pair transition due to mistake in replication caused by the presence of 2-aminopurine (AP).
(a) Nitrous acid: Nitrous acid deaminates, the bases G, C and A with decreasing frequency in DNA and with equal frequencies in RNA. Its mutagenic effect was analysed for tobacco mosaic virus (TMV), bacteria, phages T2 and T4, yeast and other organisms. HNO2 deaminates adenine (A) to hypoxanthine (H) (Fig. 23.10); cytosine (C) to uracil (U) (Fig. 23.11) and guanine (G) to xanthine (X) (Fig. 23.12). H pairs with C (Fig. 23.13); U pairs with A (Fig. 23.14) and X pairs with C (Fig. 23.15). Thus HNO2 may bring about changes like AT to GC (due to deamination of A to H); GC to AT (due to deamination of C to U) (Figs. 23.16, 23.17), but no change is brought about when guanine is deaminated to xanthine (Fig. 23.18).
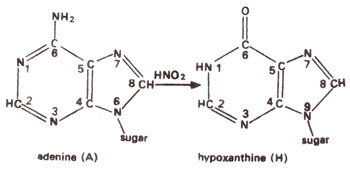
Fig. 23.10. Conversion of adenine (A) into hypoxanthine (H) due to deamination caused by nitrous acid (HNO2).
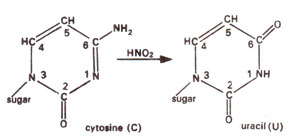
Fig. 23.11. Conversion of cytosine (C) into uracil (U) due to deamination caused by nitrous acid (HNO2).
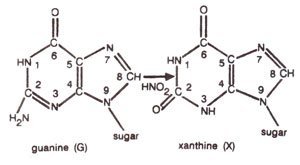
Fig. 23.12. Conversion of guanine (G) into xanthine (X) due to deamination caused by nitrous acid (HNO2).
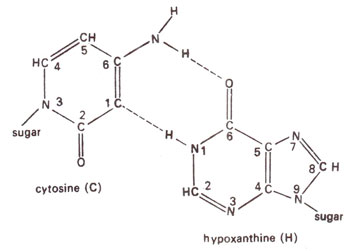
Fig. 23.13. Base pairing between cytosine (c) and hypoxanthine (H).
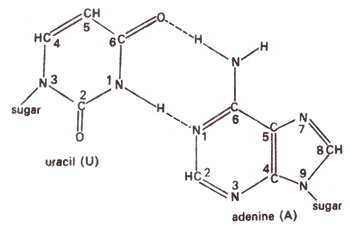
Fig. 23.14. Base pairing between adenine (A) and uracil (U).
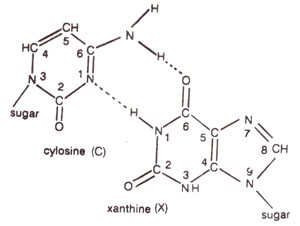
Fig. 23.15. Base pairing between cytosine (C) and xanthine (X)
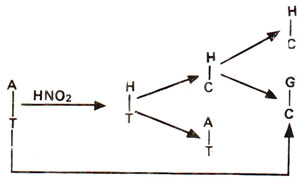
Fig. 23.16. Steps involved in AT → GC base pair transition caused due to deamination of adenine (A) into hypoxanthine (H) and H-C base pairing.
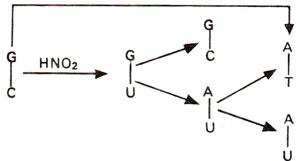
Fig. 23.17. Steps involved in GC→AT base pair transition caused due to deamination of cytosine (C) into uracil (U) followed by A-U base pairing.
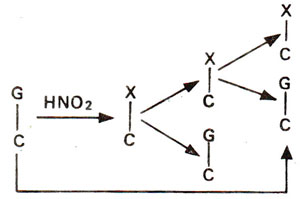
Fig. 23.18. Effect of deamination on guanine (G) to give xanthine (X) resulting into no change in G-C base pair.
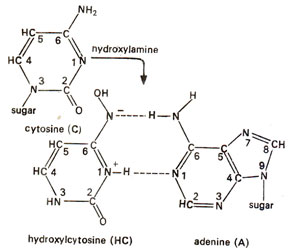
Fig. 23.19. Conversion of cytosine (C) into hydroxylcytosine (HC), which pairs with adenine (A) and not with guanine (G).
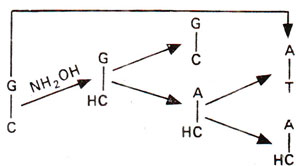
Fig. 23.20. Steps involved in GC→AT base pair transition, due to conversion of cytosine (C) into hydroxylcytosine (HC) followed by A-HC base pairing.
(c) Alkylating agents. Many mutagenic agents carry one, two or more alkyl groups in a reactive form. These are called mono-, bi- or poly functional alkylating agents. Most extensively studied alkylating agents both with tespect to their chemical effects as well as mutagenic effects are diethyl sulphate (DES), dimethyl sulphate (DMS), methyl methane sulphonate (MMS), ethylethane sulphonate (EES) and ethyl methane sulphonate (EMS). They all act as mono-functional agents even when they carry two groups like DES and DMS, since each group alkylates separately. DNA can be changed by alkylating agents in four different ways.
(ii) Hydrolysis of triester between sugar and phosphate. Hydrolysis of phosphate triester may take place between sugar and phosphate and thus causes the breakage of backbone (Fig. 23.22). This will be lethal or cause larger alterations, but will not cause point mutations.
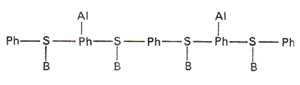
Fig. 23.21. Alkylation (Al) of phosphate (Ph) group in a polynucleotide segment of DNA.
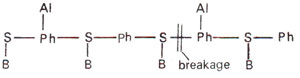
Fig. 23.22. Alkylation (Al) of phosphate (Ph) group leading to breakage of backbone of a polyniicleotide segment of DNA.
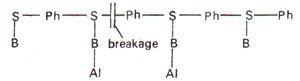
Fig. 23.23. Alkyiation (Al) of bases (B) leading to breakage of backbone of a polynucleotide segment of DNA.
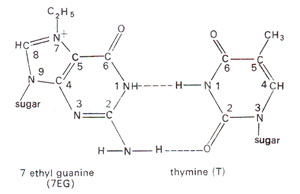
Fig. 23.24. Base pairing between 7-ethylguanine (7-EG) and thymine (T).
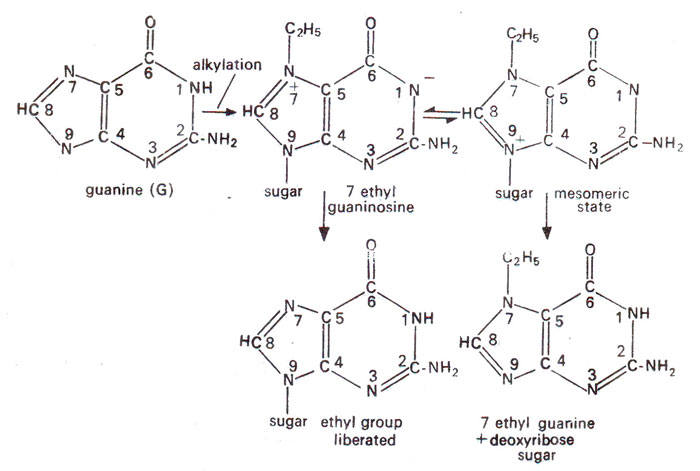
Fig. 23.25. Two different fates of 7-ethylguanine formed due to alkylation : (a) ethyl group liberated or (b) 7-ethylguanine is cleaved off (depurination).
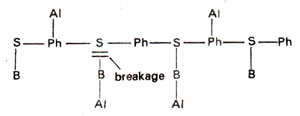
Fig. 23.26. Alkylation of phosphate (Ph) groups and bases (B) leading to breakage of sugar-base linkage.
Support our developers




