Chromosome Inversions
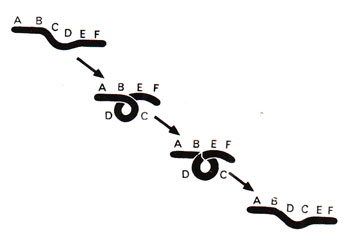
Fig. 19.15. One of the possible mechanisms, which may give rise to chromosomal inversion.
Fig. 19.15. One of the possible mechanisms, which may give rise to chromosomal inversion.
Cytology of inversions
Due to an inverted segment in one of the two homologous chromosomes, the normal kind of pairing is not possible in an inversion heterozygote. In order to enable pairing of homologous segments, a shape of loop is formed by each of the two chromosomes as shown in Figures 19.16 and 19.20. This kind of configuration will be observed both in paracentric as well as in pericentric inversions. As will be observed, the products of crossing over and the subsequent stages of meiosis will differ in these two kinds of inversions.
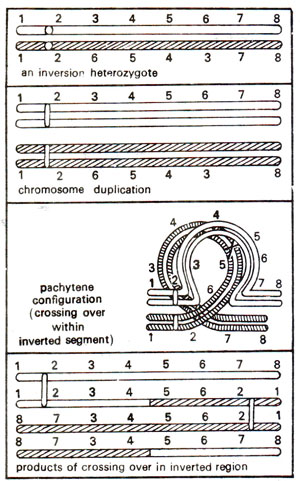
Fig. 19.16. Chromosome pairing and products of crossing over in a paracentric inversion heterozygote.
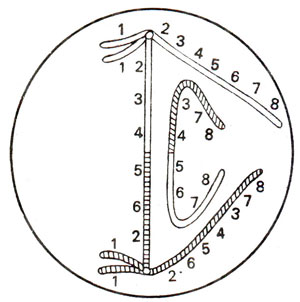
Fig. 19.17. Dicentric bridge and an acentric fragment at anaphase 1.
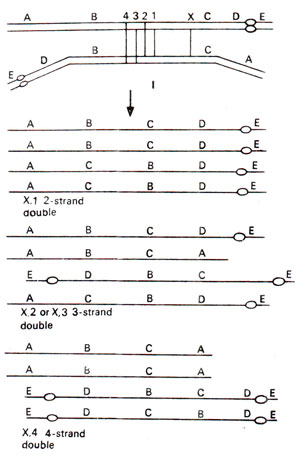
Fig. 19.18. Consequences of 2-strands, 3-strands and 4-strands double crossovers within a paracentric inversion.
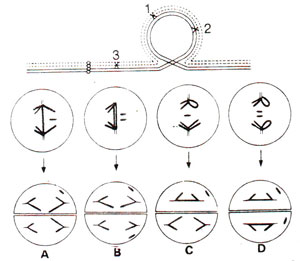
Fig. 19.19. On the top is shown inversion loop in an inversion heterozygote. Below are shown anaphase I (AI) and anaphase II (All) configurations (A-D) resulting from four different combinations of crossovers within and outside the inversion loop. A. single crossover at positions I or 2; B. 4-strand double crossovers at positions 1 and 2; C. 3-strand double crossovers, with one crossover at position 1 or 2 within inversion loop and the other crossover outside the loop at positions 3; D. 4-strand triple crossover at position 1, 2 and 3 (redrawn from McClintock, 1938).
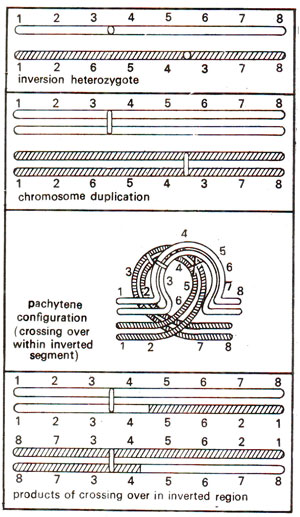
Fig. 19.20. Chromosome pairing and the products of crossing over in a pericentric inversion heterozygote.
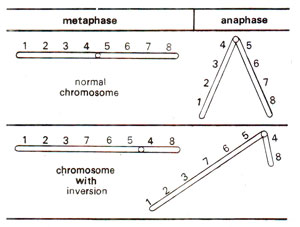
Fig. 19.21. Change in chromosome shape due to pericentric inversion.
As discussed in the preceding section on cytology, among four chromatids resulting after crossing over, the two chromatids resulting from crossing over would have deficiencies and duplications. The gametes having these chromosomes will not function. Therefore, there should be considerable gametic or zygotic lethality. In plants, there will be sufficient pollen sterility. However, since the products of single crossover will not function and the only crossovers recovered will be double crossovers, the observed frequency of recombination between any two genes in question will be considerably reduced. Due to this reason, inversions, are often called crossover suppressors. This reduction in crossing over is not the actual reduction in cytological crossing over, but is the result of lack of recovery of the products of single crossovers. This property of inversions has been utilized in the production of ClB stock, used by H.J. Mulier for the detection of sex linked lethal mutations (Mutations : 1. Morphological Level (Including Lethal Mutations)).
Sometimes a second inversion is induced in a chromosome which already has one inversion. This results in an overlapping inversion, if the segments involved in first and second inversions contain a common region. The gene orders and meiotic configurations found in an inversion heterozygote of this type are shown in Figure 19.22.
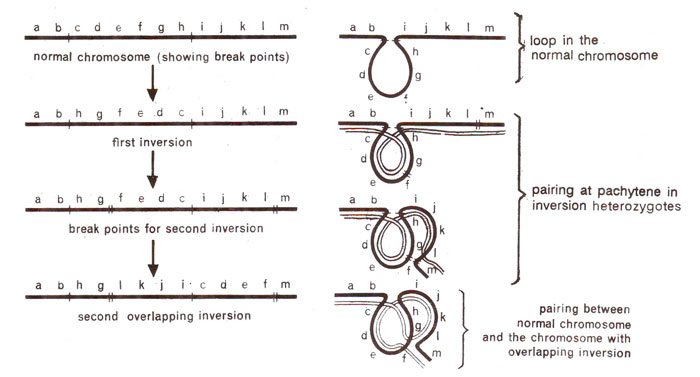
Fig. 19.22. Steps leading to overlapping inversions and corresponding pachytene configurations.
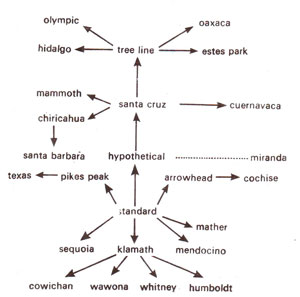
Fig. 19.23. An evolutionary tree, showing derivation of a number of races due to simple and overlapping inversions in Drosophila.
Support our developers




