Secondary growth
Content
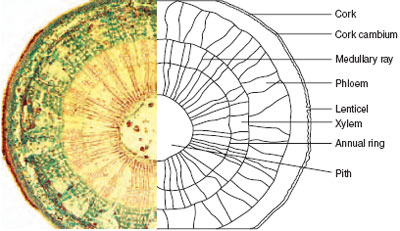
As the stem length increases, so width also increases to support the bigger plant and supply the greater amount of water and minerals required. The process in dicotyledons is called secondary growth (see Figure 6.8). Additional phloem and xylem are produced on either side of the cambium tissue, which now forms a complete ring. As these tissues increase towards the centre of the stem, so the circumference of the stem must also increase. Therefore a secondary ring of cambium (cork cambium) is formed, just to the inside of the epidermis, the cells of which divide to produce a layer of corky cells on the outside of the stem. This layer will increase with the growth of the tissue inside the stem, and will prevent loss of water if cracks should occur. As more secondary growth takes place, so more phloem and xylem tissue is produced but the phloem tubes, being soft, are squashed as the more numerous and very hard xylem vessels occupy more and more of the cross-section of the stem. Eventually, the majority of the stem consists of secondary xylem that forms the wood.
The central region of xylem sometimes becomes darkly stained with gums and resins (heartwood) and performs the long-term function of support for a heavy trunk or branch. The outer xylem, the sapwood, is still functional in transporting water and nutrients, and is often lighter in colour. The xylem tissue produced in the spring has larger diameter vessels than autumn-produced xylem, due to the greater volume of water that must be transported; a distinct ring is therefore produced where the two types of tissue meet. As these rings will be formed each season, their number can indicate the age of the branch or trunk; they are called annual rings. The phloem tissue is pushed against the cork layers by the increasing volume of xylem so that a woody stem appears to have two distinct layers, the wood in the centre and the bark on the outside. If bark is removed, the phloem also will be lost, leaving the vascular cambium exposed. The stem’s food transport system from leaves to the roots is thus removed and, if a trunk is completely ringed (or 'girdled'), the plant will die. Rabbits or deer in an orchard may cause this sort of damage. 'Partial ringing', i.e. removing the bark from almost the whole of the circumference, can achieve a deliberate reduction in growth rate of vigorous tree fruit cultivars and woody ornamental species. Initially, the bark is smooth and shiny, but with age it thickens and the outer layers accumulate chemicals (including suberin) that make it an effective protection against water loss and pest attack. This part of the bark (called cork) starts to peel or flake off. This is replaced from below and the cork gradually takes on its characteristic colours and textures. Many trees such as silver birch, London Plane, Prunus serrula, Acer davidii and many pines and rhododendrons have attractive bark and are particularly valued for winter interest (see Figure 6.9).
A further feature of a woody stem is the mass of lines radiating outwards from the centre, most obvious in the xylem tissues. These are medullary rays, consisting of parenchyma tissue linking up with small areas on the bark where the corky cells are less tightly packed together (lenticels). These allow air to move into the stem and across the stem from cell to cell in the medullary rays. The oxygen in the air is needed for the process of respiration, but the openings can be a means of entry of some diseases, e.g. Fireblight. Other external features of woody stems include the leaf scars which mark the point of attachment of leaves fallen at the end of a growing season, and can be a point of entry of fungal spores such as apple canker. Monocotyledonous stem This has the same functions as those of a dicotyledon; therefore the cell types and tissues are similar. However, the arrangement of the tissues does differ because increase in diameter by secondary growth does not take place. The stem relies on extensive sclerenchyma tissue for support that, in the maize stem shown in Figure 6.4, is found as a sheath around each of the scattered vascular bundles. Monocotyledonous stem structures are seen at their most complex in the palm family. From the outside, the trunk would appear to be made of wood, but an internal investigation shows that the stem is a mass of sclerified vascular bundles. The absence of secondary growth in the vascular bundles makes the presence of cambium tissue unnecessary. Secondary thickening is found not only in trees and shrubs, but also in many herbaceous perennials and annuals that have woody stems. However, trees and shrubs do exhibit this feature to the greatest extent. Tissues of the root
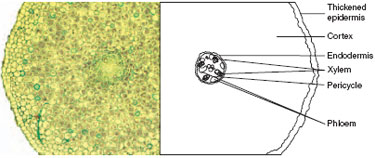
The cortex is often quite extensive and water must move across it in order to reach the transporting tissue that is in the centre of the root. This central region, called the stele, is separated from the cortex by a single layer of cells, the endodermis, which has the function of controlling the passage of water into the stele. A waxy strip forming part of the cell wall of many of the endodermal cells (the Casparian strip) prevents water from moving into the cell by all except the cells outside it, called passage cells. Water passes through the endodermis to the xylem tissue, which transports the water and dissolved minerals up to the stem and leaves. The arrangement of the xylem tissue varies between species, but often appears in transverse section as a star with varying numbers of 'arms’. Phloem tissue is responsible for transporting carbohydrates from the leaves as a food supply for the production of energy in the cortex. A distinct area in the root inside the endodermis, the pericycle, supports cell division and produces lateral roots, which push through to the main root surface from deep within the structure. Roots age and become thickened with waxy substances, and the uptake rate of water becomes restricted. |
||||||||||||||||||||||||||


Support our developers




