Other Amphibia
Other AmphibiaIn all the Amphibia, a glottis, placed on the ventral wall of the oesophagus, opens into a short laryngo-tracheal chamber with which two pulmonary sacs are connected, either directly, or by the intermediation of bronchi (as in the Aglossa), or by a trachea (as in the Gymnophiona). The walls of the pulmonary sacs are more or less sacculated. In most Amphibia the lungs are equal in size; but in the snake-like Gymnophiona, the right is much smaller than the left. In Proteus, the pulmonary blood is not all returned to the heart, some of it entering the veins of the trunk. Aerial respiration is effected, in the Amphibia, by pumping the air from the oral cavity into the lungs. To this end the mouth is kept Shut, and ingress and egress to the air is given by the nasal passages, which always open immidiately behind the vomers, at the anterior part of the roof of the mouth. These passages being open, and the hyoidean apparatus depressed, the air fills the cavity of the mouth. The external nortils are than shut, and, the hyoidean apparatus being raised, the air is forced, through the open glottis, into the lungs.
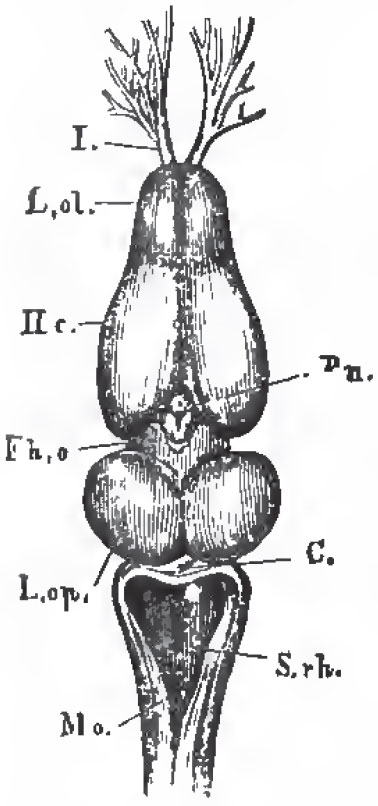 |
| Fig. 59. - The brain of Rana esculenta from above, magnified four times; L.ol.,the rhinencephalon, or olfactory jobes, with, I., the olfactory nerves; Hc., the cerebral hemispheres; Fh. o., the thalamencephalon with the pineal glaud, Pn., L. op., optic lobes; C., cerebellum; S. rh., the fourth ventricle; Mo., medulla oblongata. |
In the brain of the Amphibia the cerebellum is always very small, and represented by a mere band; the cerebral hemispheres are elongated, and contain ventricles. In Proteus the mesencephalon is very indistinctly marked. The optic nerves form a chiasma.
As in fishes, the pneumogastric gives off a lateral nerve, which runs along the sides of the body.
The eyes are very small, and covered by the integument, in Proteus, the Gymnophiona, and the genus Pipa. The perennibranchiate and derotreme Urodela have no eyelids; but most Batrachia have not only a well-developed upper eyelid, but a nictitating membrane, moved by special muscles.
All Amphibia possess a fenestra ovalis with a cartilaginous, or osseous, columelliform stapes, the expanded proximal end of which is fixed to the membrane of the fenestra. In many Batrachia, if not in all, there is a fenestra rotunda, though the presence of a distinct cochlea has not been ascertained. The Urodela, the Gymnophiona, and the Pelobatidea, among the Batrachia, have no tympanic cavity, nor membrane. In the other Batrachia there are tympanic cavities communicating freely with the throat. Each is closed externally by a tympanic membrane, with which the outer extremity of the stem of the stapes is connected. In the Aglossa, the two tympanic cavities communicate with the mouth by a single Eustachian aperture; and the outer end of the stapes expands into a great cartilaginous plate coextensive with the tympanic membrane.
The ducts of the reproductive organs of the Amphibia, like those of the Ganoidei, always communicate directly with the urinary ducts: and, as in most Ganoidei and all Elasmohranchii, the proximal end of the oviduct is open, and communicates with the peritoneal cavity. The male has no penis, unless a papillary elevation of the wall of the cloaca may represent such an organ. The testes of the male Amphibia are composed of tubules, and vasa efferentia convey the contents of these away. In the Urodela, the vasa efferentia of each testis enter the inner side of the corresponding kidney, and traverse it, leaving its outer side to enter a genito-urinary duct, which lies on the outer side of the kidney, ends blindly in front, and opens behind into the cloaca. The uriniferous tubuli also pass directly from the outer margin of the kidneys into the genito-urinary duct. In the Batrachia there is likewise a genito-urinary duct, and the vasa efferentia run to the inner edge of the kidney and enter it. In Bombinator igneua and Discoglossus pictus, the genito-urinary duct receives the urinary products and the spermatozoa, in the same way as in the Urodela. But, in the Frogs and Toads, the urinary tubuli are gathered together into a special small canal which opens into the genito-urinary duct near its termination in the cloaca, and the vasa efferentia pour their contents into this canal. Under these circumstances, the part of the genito-urinary duct which lies beyond the renal canal may become obliterated, as in the Frogs; or may persist, and play the part of a vesicula seminalis, as in the Toads.
In the female Amphibia, the kidneys have, as in the male Frogs and Toads, a renal canal which opens into the lower part of the oviduct.
It would appear from these facts that the oviduct in the female, and the genito-urinary ducts in the male, Amphibia represent both the Wolffian and the Mullerian ducts of the higher Vertebrata.
In most Amphibia the ova are impregnated and hatched outside the body, but internal impregnation and incubation occur in some of the Urodela. In Pipa the eggs are hatched in pouches of the dorsal integument, while the male Alytes carries them twisted in strings round his legs.
When hatched, the young are devoid of respiratory organs and of limbs, and are provided with a long tail, by means of which they swim about. Branchial clefts soon make their appearance; and ciliated external branchial plumes, like those of the perennibranchiate Urodela, are developed. A pair of suckers are sometimes formed upon the under-surface of the mandibular region, and the jaws acquire horny sheaths.
A broad opercular membrane is developed in front of the branchial aperture, and, in the Batrachia, extends over and eventually covers the gills, a rounded aperture persisting for a certain time only on the left side. The anterior pair of limbs is developed before the posterior, but in the Frog they are not so soon visible, being hidden by the opercular membrane.
The lungs make their appearance as diverticula of the ventral wall of the oesophagus. The nasal sacs are at first mere caecal involutions of the integument, but nasal passages communicating with the mouth are soon formed, and both aerial and aquatic respiration are completely established.
In the Batrachia, as development proceeds, the external branchiae disappear, and are succeeded, functionally, by short branchial filaments developed upon the whole length of each of the branchial arches, of which there are four.
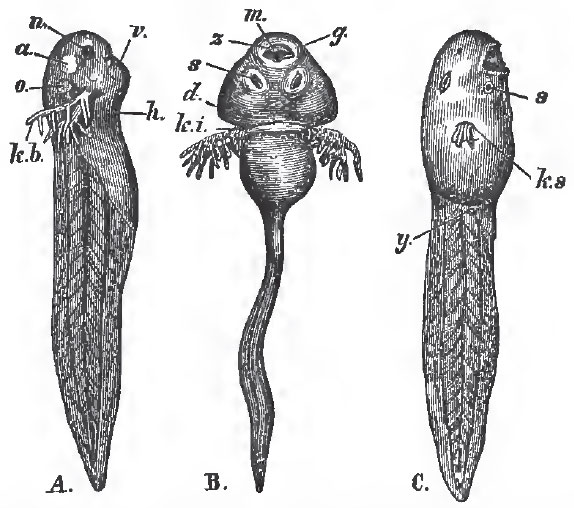 |
| Fig. 60. - A. B. Tadpoles with external branchlae: n, nasal sacs: a, eye; o, ear; k b, branchiae; m, mouth; z, horny jaws; s, suckers; d, opercular fold. C., a more advanced Frog's larva: y, the rudiment of tne hind-limb; k, s, the single branchial aperture. The figure has not been reversed, so that this aperture appears to lie on the nght side instead of the left. |
Before the development of the lungs the heart has only a single auricle; afterward, the auricle becomes divided into two. The aortic arches, at first, pass along the visceral and branchial arches to the dorsal aorta, as in other vertebrate embryos. When external gills are developed, each receives a loop from the corresponding arch, much as in Proteus.
When the internal gills of the Batrachia appear, each aortic arch which belongs to a branchial arch splits into two trunks, - one which remains directly connected with the cardiac aorta, and another which opens into the dorsal aorta. The vessels of the branchial filaments constitute loops between these afferent and efferent trunks, which always remain united by anastomoses. When branchial respiration ceases, and the branchial processes and their vessels disappear, the anastomoses dilate; the direct communication between the afferent and efferent trunks of the second pair of internal branchiae ia reestablished; and they become the permanent arches of the aorta. The anterior branchiae are replaced by the carotid glands, and their afferent vessel is the carotid passage of the adult. The afferent and efferent trunks of the third pair of branchiae are converted into the stem of the cutaneous artery, and the afferent trunk of the fourth pair of branchiae into that of the pulmonary artery. The diagram (Fig. 25, p. 83) is intended to make these changes, and the relations of the various trunks to the embryonic aortic arches, intelligible.
The alimentary canal of the Tadpole is, at first, long, and coiled up into a close spire, like a watch-spring, in the abdomen, but its length becomes relatively less as age advances. At the same time, the diet changes from vegetable to animal-the young tadpole being chiefly herbivorous, the adult, insectivorous.
Support our developers




