Human Evolution
Human Evolution
Darwin devoted an entire book, The Descent of Man and Selection in Relation to Sex (1871), largely to human evolution. The idea that humans shared common descent with apes and other animals was repugnant to the Victorian world, which responded with predictable outrage (Figure 6-14). Because at that time virtually no fossil evidence linking humans with apes existed, Darwin built his case mostly on anatomical comparisons between humans and apes. To Darwin, the close resemblances between apes and humans could be explained only by common descent.
The search for fossils, especially for a “missing link” that would provide a connection between apes and humans, began when two skeletons of Neanderthals were collected in the 1880s. Then in 1891, Eugene Dubois discovered the famous Java man (Homo erectus). The most spectacular discoveries, however, have been made in Africa, especially between 1967 and 1977, which American paleoanthropologist Donald C. Johanson calls the “golden decade.” During this same period, comparative biochemical studies demonstrated humans and chimpanzees are as similar genetically as many sibling species. Comparative cytology provided evidence that chromosomes of humans and apes are homologous. We are no longer searching for a mythical “missing link” to establish the common descent of humans and apes, our closest living relatives.
Evolutionary Radiation of the Primates
Humans are primates, a fact that even the pre-evolutionist Linnaeus recognized. All primates share certain significant characteristics: grasping fingers on all four limbs, flat fingernails instead of claws, and forward-pointing eyes with binocular vision and excellent depth perception. Details of primate phylogeny are not entirely clear. The following synopsis will highlight the probable relationships of the major primate groups.
The earliest primate was probably a small, nocturnal animal similar in appearance to tree shrews. This ancestral primate stock split into two major lineages, one of which gave origin to the prosimians, including lemurs, tarsiers (Figure 30-30), and lorises; and the other to the simians, which include the monkeys (Figure 30-31) and apes (Figure 30-32). Prosimians and many simians are arboreal (tree-dwellers), which is probably the ancestral life-style for both groups. Arboreality probably selected for increased intelligence. Flexible limbs are essential for active animals moving through trees. Grasping hands and feet, in contrast to the clawed feet of squirrels and other rodents, enable primates to grip limbs, hang from branches, seize and manipulate food, and, most significantly, use tools. Highly developed sense organs, especially good forward and peripheral vision, and proper coordination of limb and finger muscles are essential for an active arboreal life. Of course, sense organs are no better than the brain processing sensory information. Precise timing, judgment of distance, and alertness require a large cerebral cortex.
The earliest simian fossils appeared
in Africa in late Eocene
deposits, some 40 million years ago. Many of these primates became diurnal
rather than nocturnal, making vision
the dominant special sense, now enhanced
by color vision. We recognize
three major simian groups whose precise
phylogenetic relationships are
unknown. These are (1) the New
World monkeys of South America
(ceboids; Figure 30-31A), including the
howler monkey, spider monkey, and
the tamarin, (2) the Old World monkeys
(cercopithecoids), including the
baboon (Figure 30-31B), mandrill, and
the colobus monkey, and (3) the
anthropoid apes (Figure 30-32). Old
World monkeys and anthropoid apes
(including humans) are sister taxa, and
together form the sister group of the
New World monkeys. In addition to
their geographic separation, Old World
monkeys differ from New World monkeys
in lacking a grasping tail, while
having close-set nostrils, better opposable,
grasping thumbs, and more
advanced teeth. Apes first appear in
25-million-year-old fossils. At this time
the woodland savannas were arising in
Africa, Europe, and North America.
Perhaps motivated by the greater
abundance of food on the ground,
these apes left the trees and became
largely terrestrial.
The First Hominids
About 8 million years ago during the Miocene epoch the gradual replacement of forests with grasslands in eastern Africa provided an impetus for apes to adapt to an open environment, the savannas. Because of the benefits of standing upright (better view of predators, freeing of hands for using tools, defense, caring for young, and gathering food) emerging hominids gradually evolved upright posture. This important transition was an enormous leap because it required extensive redesigning of the skeleton and muscle attachments.
Evidence of the earliest hominids of this period is remarkably sparse. Although several different early hominids have been identified, they all disappeared without a trace of their descendants. Not until about 4.4 million years ago, after a lengthy fossil gap, do the first “near humans” appear. One was Australopithecus afarensis, a short, bipedal hominid with a face and brain size resembling those of a chimpanzee. Numerous fossils of this species have now been unearthed, the most celebrated of which was the 40% complete skeleton of a female discovered in 1974 by Donald Johanson and named “Lucy” (Figures 30-33 and 30-34). The time range for A. afarensis is 3.7 to 3.0 million years ago. Since the discovery of Lucy new finds of even older hominid fossils have extended our knowledge of human ancestry. In 1994 the sands of Ethiopia yielded teeth and bone fragments of the most ancient hominid yet discovered, Ardipithecus ramidus, dated at 4.4 million years ago. A. ramidus, a mosaic of primitive apelike and derived hominid traits, appears to be ancestral to the australopithecine species. In 1995 Australopithecus anamensis was discovered in the Rift Valley of Kenya. Many researchers agree that this species, which may have been bipedal, is an intermediate between A. ramidus and A. afarensis (Lucy). A. anamensis lived between 4.2 to 3.9 million years ago.
Between 3 and 4 million years ago two quite separate hominid lines emerged that coexisted for at least 2 million years. One lineage was Homo, our own genus, described below. The other was the bipedal australopithecine lineage. Australopithecus africanus, which lived between 2.8 and 2.3 million ago, had a more humanlike face than A. afarensis and a brain size about one-third as large as that of modern humans. A different line of large and robust australopithecines included Paranthropus robustus (Figure 30-35), which probably approached the size of a gorilla. The “robust” australopithecines were heavy jawed with skull crests and large back molars, perhaps for eating roots and tubers. They are a side branch in hominid evolution and not part of our own lineage. In 1998 Australopithecus garhi, dated at 2.5 million years ago, was discovered in Ethiopia. This most recent fossil appears to be a descendant of A. afarensis and thus a possible candidate for an ancestor for Homo.
Emergence of Homo, the True Human
Although researchers are deeply divided over who the first members of Homo were, and indeed how to define the genus Homo, it is generally agreed that at least three species of Homo shared the African landscape with the australopithecines. One of these was Homo habilis (Figure 30-35), a fully erect hominid, more lightly built and larger brained than the australopithecines. Homo habilis unquestionably used stone and bone tools. This species appeared about 2 million years ago and survived for perhaps 500,000 years.
About 1.5 million years ago Homo erectus appeared, a large hominid standing 150 to 170 cm (5 to 5.5 feet) tall, with a low but distinct forehead and strong browridges. The brain capacity was around 1000 cc, about intermediate between the brain capacity of Homo habilis and modern humans (Figure 30-35). Homo erectus was a social species living in tribes of 20 to 50 individuals. Homo erectus had a successful and complex culture and became widespread throughout the tropical and temperate Old World.
Homo Sapiens: Modern Hominids
After the disappearance of Homo erectus around 300,000 years ago, subsequent human evolution and the establishment of Homo sapiens (“wise man”) threaded a complex course. From among many early subcultures of Homo sapiens, the Neanderthals emerged about 130,000 years ago (Figure 30-35). With a brain capacity well within range of modern humans, the Neanderthals were proficient hunters and tool-users. Neanderthals were not homogeneous but varied geographically in response to local conditions and the isolation of populations from one another. They dominated the Old World in the late Pleistocene epoch.
About 30,000 years ago Neanderthals were replaced and quite possibly exterminated by modern humans. The geographical origin of modern humans is obscure. They were tall people with a culture very different from that of Neanderthals. Implement crafting developed rapidly, and human culture became enriched with aesthetics, artistry, and sophisticated language.
In closing our discussion of human evolution, it is important to note that recognition of species in Homo is based entirely on morphology. Recognition of three or more distinct species of Homo does not necessarily imply the occurrence of branching speciation in this lineage; it is perhaps equally likely that we are observing phyletic change within a single species through time, and using the species names only to denote different grades of evolution. Nevertheless, there is clearly only a single species of Homo alive today.
The Unique Human Position
Biologically, Homo sapiens is a product of the same processes that have directed the evolution of every organism from the time of life’s origin. Mutation, isolation, genetic drift, and natural selection have operated for us as they have for other animals. Yet we have what no other animals has, a nongenetic cultural evolution that provides a constant feedback between past and future experience. Our symbolic languages, capacities for conceptual thought, knowledge of our history, and abilities to manipulate our environment emerge from this nongenetic cultural endowment. Finally, we owe much of our cultural and intellectual achievements to our arboreal ancestry which bequeathed us with binocular vision, superb visuotactile discrimination, and manipulative skills in the use of our hands. If the horse (with one toe instead of five fingers) had human mental capacity, could it have accomplished what humans have?
Darwin devoted an entire book, The Descent of Man and Selection in Relation to Sex (1871), largely to human evolution. The idea that humans shared common descent with apes and other animals was repugnant to the Victorian world, which responded with predictable outrage (Figure 6-14). Because at that time virtually no fossil evidence linking humans with apes existed, Darwin built his case mostly on anatomical comparisons between humans and apes. To Darwin, the close resemblances between apes and humans could be explained only by common descent.
 |
| Figure 30-30 A prosimian, the Mindanao tarsier, Tarsius syrichta carbonarius of Mindanao Island in the Philippines. Order Primates, family Tarsiidae. |
The search for fossils, especially for a “missing link” that would provide a connection between apes and humans, began when two skeletons of Neanderthals were collected in the 1880s. Then in 1891, Eugene Dubois discovered the famous Java man (Homo erectus). The most spectacular discoveries, however, have been made in Africa, especially between 1967 and 1977, which American paleoanthropologist Donald C. Johanson calls the “golden decade.” During this same period, comparative biochemical studies demonstrated humans and chimpanzees are as similar genetically as many sibling species. Comparative cytology provided evidence that chromosomes of humans and apes are homologous. We are no longer searching for a mythical “missing link” to establish the common descent of humans and apes, our closest living relatives.
Evolutionary Radiation of the Primates
Humans are primates, a fact that even the pre-evolutionist Linnaeus recognized. All primates share certain significant characteristics: grasping fingers on all four limbs, flat fingernails instead of claws, and forward-pointing eyes with binocular vision and excellent depth perception. Details of primate phylogeny are not entirely clear. The following synopsis will highlight the probable relationships of the major primate groups.
The earliest primate was probably a small, nocturnal animal similar in appearance to tree shrews. This ancestral primate stock split into two major lineages, one of which gave origin to the prosimians, including lemurs, tarsiers (Figure 30-30), and lorises; and the other to the simians, which include the monkeys (Figure 30-31) and apes (Figure 30-32). Prosimians and many simians are arboreal (tree-dwellers), which is probably the ancestral life-style for both groups. Arboreality probably selected for increased intelligence. Flexible limbs are essential for active animals moving through trees. Grasping hands and feet, in contrast to the clawed feet of squirrels and other rodents, enable primates to grip limbs, hang from branches, seize and manipulate food, and, most significantly, use tools. Highly developed sense organs, especially good forward and peripheral vision, and proper coordination of limb and finger muscles are essential for an active arboreal life. Of course, sense organs are no better than the brain processing sensory information. Precise timing, judgment of distance, and alertness require a large cerebral cortex.
 |
| Figure 30-31 Monkeys. A, Red-howler monkeys, Order Primates, family Cebidae, an example of the New World monkeys. B, The olive baboon, order Primates, family Cercopithecidae, an example of the Old World monkeys |
 |
| Figure 30-32 The gorilla, order Primates, family Hominidae, an example of the anthropoid apes. |
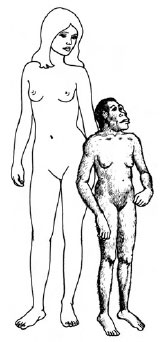 |
| Figure 30-34 A reconstruction of the appearance of Lucy (right) compared with a modern human (Homo sapiens) order Primates (left). |
The First Hominids
About 8 million years ago during the Miocene epoch the gradual replacement of forests with grasslands in eastern Africa provided an impetus for apes to adapt to an open environment, the savannas. Because of the benefits of standing upright (better view of predators, freeing of hands for using tools, defense, caring for young, and gathering food) emerging hominids gradually evolved upright posture. This important transition was an enormous leap because it required extensive redesigning of the skeleton and muscle attachments.
Evidence of the earliest hominids of this period is remarkably sparse. Although several different early hominids have been identified, they all disappeared without a trace of their descendants. Not until about 4.4 million years ago, after a lengthy fossil gap, do the first “near humans” appear. One was Australopithecus afarensis, a short, bipedal hominid with a face and brain size resembling those of a chimpanzee. Numerous fossils of this species have now been unearthed, the most celebrated of which was the 40% complete skeleton of a female discovered in 1974 by Donald Johanson and named “Lucy” (Figures 30-33 and 30-34). The time range for A. afarensis is 3.7 to 3.0 million years ago. Since the discovery of Lucy new finds of even older hominid fossils have extended our knowledge of human ancestry. In 1994 the sands of Ethiopia yielded teeth and bone fragments of the most ancient hominid yet discovered, Ardipithecus ramidus, dated at 4.4 million years ago. A. ramidus, a mosaic of primitive apelike and derived hominid traits, appears to be ancestral to the australopithecine species. In 1995 Australopithecus anamensis was discovered in the Rift Valley of Kenya. Many researchers agree that this species, which may have been bipedal, is an intermediate between A. ramidus and A. afarensis (Lucy). A. anamensis lived between 4.2 to 3.9 million years ago.
 |
| Figure 30-33
Lucy (Australopithecus afarensis), the most nearly complete skeleton of an early hominid ever found. Lucy is dated at 2.9 million years old. A nearly complete skull of A. afarensis was discovered in 1994. |
Between 3 and 4 million years ago two quite separate hominid lines emerged that coexisted for at least 2 million years. One lineage was Homo, our own genus, described below. The other was the bipedal australopithecine lineage. Australopithecus africanus, which lived between 2.8 and 2.3 million ago, had a more humanlike face than A. afarensis and a brain size about one-third as large as that of modern humans. A different line of large and robust australopithecines included Paranthropus robustus (Figure 30-35), which probably approached the size of a gorilla. The “robust” australopithecines were heavy jawed with skull crests and large back molars, perhaps for eating roots and tubers. They are a side branch in hominid evolution and not part of our own lineage. In 1998 Australopithecus garhi, dated at 2.5 million years ago, was discovered in Ethiopia. This most recent fossil appears to be a descendant of A. afarensis and thus a possible candidate for an ancestor for Homo.
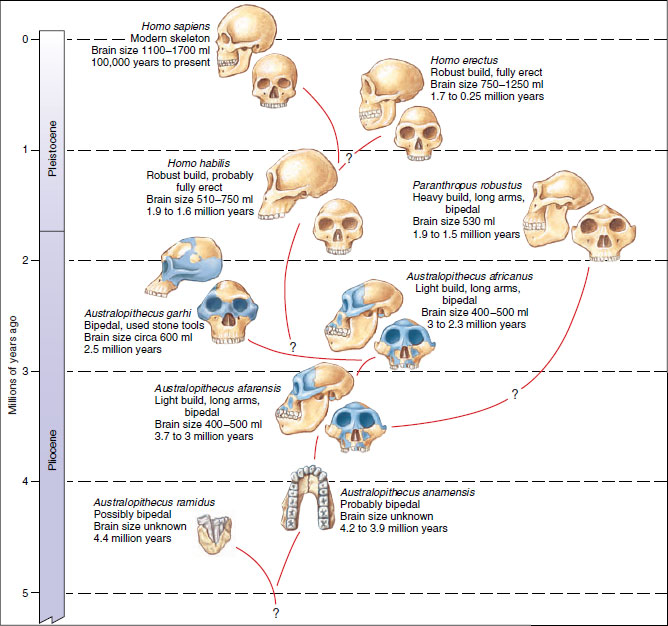 |
| Figure 30-35 Hominid skulls, showing several of the best-known hominid lines preceding modern humans (Homo sapiens). The time span of existence for each species, as indicated by the fossil record, is suggested by the vertical red lines. |
Emergence of Homo, the True Human
Although researchers are deeply divided over who the first members of Homo were, and indeed how to define the genus Homo, it is generally agreed that at least three species of Homo shared the African landscape with the australopithecines. One of these was Homo habilis (Figure 30-35), a fully erect hominid, more lightly built and larger brained than the australopithecines. Homo habilis unquestionably used stone and bone tools. This species appeared about 2 million years ago and survived for perhaps 500,000 years.
About 1.5 million years ago Homo erectus appeared, a large hominid standing 150 to 170 cm (5 to 5.5 feet) tall, with a low but distinct forehead and strong browridges. The brain capacity was around 1000 cc, about intermediate between the brain capacity of Homo habilis and modern humans (Figure 30-35). Homo erectus was a social species living in tribes of 20 to 50 individuals. Homo erectus had a successful and complex culture and became widespread throughout the tropical and temperate Old World.
Homo Sapiens: Modern Hominids
After the disappearance of Homo erectus around 300,000 years ago, subsequent human evolution and the establishment of Homo sapiens (“wise man”) threaded a complex course. From among many early subcultures of Homo sapiens, the Neanderthals emerged about 130,000 years ago (Figure 30-35). With a brain capacity well within range of modern humans, the Neanderthals were proficient hunters and tool-users. Neanderthals were not homogeneous but varied geographically in response to local conditions and the isolation of populations from one another. They dominated the Old World in the late Pleistocene epoch.
About 30,000 years ago Neanderthals were replaced and quite possibly exterminated by modern humans. The geographical origin of modern humans is obscure. They were tall people with a culture very different from that of Neanderthals. Implement crafting developed rapidly, and human culture became enriched with aesthetics, artistry, and sophisticated language.
In closing our discussion of human evolution, it is important to note that recognition of species in Homo is based entirely on morphology. Recognition of three or more distinct species of Homo does not necessarily imply the occurrence of branching speciation in this lineage; it is perhaps equally likely that we are observing phyletic change within a single species through time, and using the species names only to denote different grades of evolution. Nevertheless, there is clearly only a single species of Homo alive today.
The Unique Human Position
Biologically, Homo sapiens is a product of the same processes that have directed the evolution of every organism from the time of life’s origin. Mutation, isolation, genetic drift, and natural selection have operated for us as they have for other animals. Yet we have what no other animals has, a nongenetic cultural evolution that provides a constant feedback between past and future experience. Our symbolic languages, capacities for conceptual thought, knowledge of our history, and abilities to manipulate our environment emerge from this nongenetic cultural endowment. Finally, we owe much of our cultural and intellectual achievements to our arboreal ancestry which bequeathed us with binocular vision, superb visuotactile discrimination, and manipulative skills in the use of our hands. If the horse (with one toe instead of five fingers) had human mental capacity, could it have accomplished what humans have?
Support our developers




