Origin and Adaptive Radiation of Reptilian Groups
Origin and Adaptive
Radiation of
Reptilian Groups
As mentioned in the prologue to this section, amniotes are a monophyletic group that evolved in the late Paleozoic. Most paleontologists agree that the amniotes arose from a group of amphibian-like tetrapods, the anthracosaurs, during the early Carboniferous period of the Paleozoic. By the late Carboniferous (approximately 300 million years ago), amniotes had separated into three lineages. The first lineage, the anapsids (Gr. an, without, + apsis, arch), is characterized by a skull having no temporal opening behind the orbits, the skull behind the orbits being completely roofed with dermal bone (Figure 28-2). This group is represented today only by the turtles. Their morphology is an odd mix of ancestral and derived characters that has scarcely changed at all since turtles first appeared in the fossil record in the Triassic some 200 million years ago.
The second lineage, the diapsids (Gr. di, double, + apsis, arch), gave rise to all other reptilian groups and to the birds (Figure 28-1). The diapsid skull was characterized by the presence of two temporal openings: one pair located low on the cheeks, and a second pair positioned above the lower pair and separated from them by a bony arch (Figure 28-2). Three subgroups of diapsids appeared. The lepidosaurs include extinct marine ichthyosaurs and all modern reptiles except the turtles and crocodilians. The more derived archosaurs comprised dinosaurs and their relatives, and living crocodilians and birds. A third, smaller subgroup, the sauropterygians included several extinct aquatic groups, most conspicuous of which were the large, long-necked plesiosaurs.
The third lineage was the synapsids (Gr. syn, together, + apsis, arch), the mammal-like reptiles. The synapsid skull had a single pair of temporal openings located low on the cheeks and bordered by a bony arch (Figure 28-2). The synapsids were the first amniote group to diversify, giving rise first to pelycosaurs, later to therapsids, and finally to mammals (Figure 28-1).
Changes in Traditional Classification of Reptilian Groups
With increasing use of cladistic methodology in zoology, and its insistence on hierarchical arrangement of monophyletic groups, important changes have been made in the traditional classification of reptiles. The class Reptilia is no longer recognized by cladists as a valid taxon because it is not monophyletic. As customarily defined, class Reptilia excludes birds, which descend from the most recent common ancestor of the reptiles. Consequently, reptiles are a paraphyletic group because they do not include all descendants of their most recent common ancestor. Reptiles can be identified only as amniotes that are not birds. This is clearly shown in the phylogenetic tree of the amniotes (Figure 28-1).
An example of this problem is the shared ancestry of birds and crocodilians. Based solely on shared derived characteristics, crocodilians and birds are sister groups; they are more recently descended from a common ancestor than either is from any other living reptilian lineage. In other words, birds and crocodilians belong to a monophyletic group apart from other reptiles and, according to the rules of cladism, should be assigned to a clade that separates them from the remaining reptiles. This clade is in fact recognized; it is the Archosauria (Figures 28-1 and 28-2), a grouping that also includes the extinct dinosaurs. Therefore birds should be classified as reptiles. Archosaurs plus their sister group, the lepidosaurs (tuataras, lizards, snakes, and amphisbaenids), comprise a monophyletic group that some taxonomists call the Reptilia. The term “Reptilia” is thereby redefined to include birds in contrast to its traditional usage. However, evolutionary taxonomists argue that birds represent a novel adaptive zone and grade of organization whereas crocodilians remain within the traditionally recognized reptilian adaptive zone and grade. In this view, the morphological and ecological novelty of birds has been recognized by maintaining the traditional classification that places crocodilians in class Reptilia and birds in class Aves. Such conflicts of opinion between proponents of the two major competing schools of taxonomy (cladistics and evolutionary taxonomy) have had the healthy effect of forcing zoologists to reevaluate their views of amniote genealogy and how vertebrate classifications should represent genealogy and degrees of divergence. In our treatment, “reptilian group,” “reptile,” and “reptilian” refer to members of four monophyletic groups (turtles, crocodilians, squamates, tuataras) that formerly were combined into the paraphyletic class Reptilia.
Characteristics of Class Reptilia
As mentioned in the prologue to this section, amniotes are a monophyletic group that evolved in the late Paleozoic. Most paleontologists agree that the amniotes arose from a group of amphibian-like tetrapods, the anthracosaurs, during the early Carboniferous period of the Paleozoic. By the late Carboniferous (approximately 300 million years ago), amniotes had separated into three lineages. The first lineage, the anapsids (Gr. an, without, + apsis, arch), is characterized by a skull having no temporal opening behind the orbits, the skull behind the orbits being completely roofed with dermal bone (Figure 28-2). This group is represented today only by the turtles. Their morphology is an odd mix of ancestral and derived characters that has scarcely changed at all since turtles first appeared in the fossil record in the Triassic some 200 million years ago.
The second lineage, the diapsids (Gr. di, double, + apsis, arch), gave rise to all other reptilian groups and to the birds (Figure 28-1). The diapsid skull was characterized by the presence of two temporal openings: one pair located low on the cheeks, and a second pair positioned above the lower pair and separated from them by a bony arch (Figure 28-2). Three subgroups of diapsids appeared. The lepidosaurs include extinct marine ichthyosaurs and all modern reptiles except the turtles and crocodilians. The more derived archosaurs comprised dinosaurs and their relatives, and living crocodilians and birds. A third, smaller subgroup, the sauropterygians included several extinct aquatic groups, most conspicuous of which were the large, long-necked plesiosaurs.
The third lineage was the synapsids (Gr. syn, together, + apsis, arch), the mammal-like reptiles. The synapsid skull had a single pair of temporal openings located low on the cheeks and bordered by a bony arch (Figure 28-2). The synapsids were the first amniote group to diversify, giving rise first to pelycosaurs, later to therapsids, and finally to mammals (Figure 28-1).
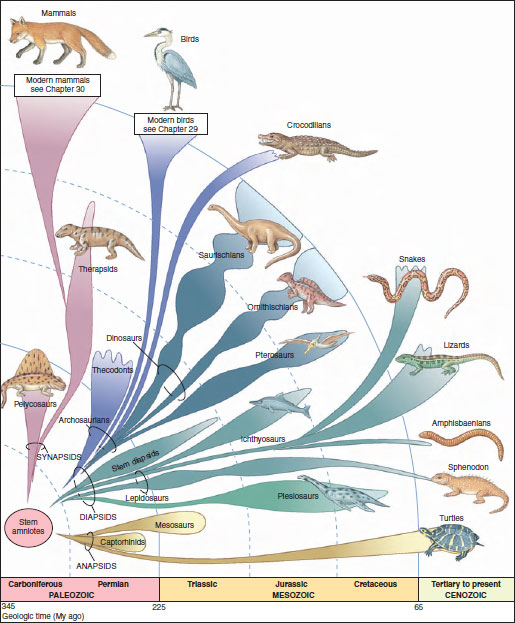 |
| Figure 28-1 Evolution of the amniotes. The evolutionary origin of amniotes occurred by the evolution of an amniotic egg that made reproduction on land possible, although this egg may well have developed before the earliest amniotes had ventured far on land. The amniote assemblage, which includes the reptiles, birds, and mammals, evolved from a lineage of small, lizardlike forms known as captorhinids that retained the skull pattern of the early tetrapods. First to diverge from the primitive stock were the mammal-like reptiles, characterized by a skull pattern termed the synapsid condition. All other amniotes, including the birds and all living reptiles except the turtles, have a skull pattern known as diapsid. The turtles have a skull pattern known as anapsid. The great Mesozoic radiation of reptiles may have resulted partly from the increased variety of ecological habitats into which the amniotes could move. |
Changes in Traditional Classification of Reptilian Groups
With increasing use of cladistic methodology in zoology, and its insistence on hierarchical arrangement of monophyletic groups, important changes have been made in the traditional classification of reptiles. The class Reptilia is no longer recognized by cladists as a valid taxon because it is not monophyletic. As customarily defined, class Reptilia excludes birds, which descend from the most recent common ancestor of the reptiles. Consequently, reptiles are a paraphyletic group because they do not include all descendants of their most recent common ancestor. Reptiles can be identified only as amniotes that are not birds. This is clearly shown in the phylogenetic tree of the amniotes (Figure 28-1).
An example of this problem is the shared ancestry of birds and crocodilians. Based solely on shared derived characteristics, crocodilians and birds are sister groups; they are more recently descended from a common ancestor than either is from any other living reptilian lineage. In other words, birds and crocodilians belong to a monophyletic group apart from other reptiles and, according to the rules of cladism, should be assigned to a clade that separates them from the remaining reptiles. This clade is in fact recognized; it is the Archosauria (Figures 28-1 and 28-2), a grouping that also includes the extinct dinosaurs. Therefore birds should be classified as reptiles. Archosaurs plus their sister group, the lepidosaurs (tuataras, lizards, snakes, and amphisbaenids), comprise a monophyletic group that some taxonomists call the Reptilia. The term “Reptilia” is thereby redefined to include birds in contrast to its traditional usage. However, evolutionary taxonomists argue that birds represent a novel adaptive zone and grade of organization whereas crocodilians remain within the traditionally recognized reptilian adaptive zone and grade. In this view, the morphological and ecological novelty of birds has been recognized by maintaining the traditional classification that places crocodilians in class Reptilia and birds in class Aves. Such conflicts of opinion between proponents of the two major competing schools of taxonomy (cladistics and evolutionary taxonomy) have had the healthy effect of forcing zoologists to reevaluate their views of amniote genealogy and how vertebrate classifications should represent genealogy and degrees of divergence. In our treatment, “reptilian group,” “reptile,” and “reptilian” refer to members of four monophyletic groups (turtles, crocodilians, squamates, tuataras) that formerly were combined into the paraphyletic class Reptilia.
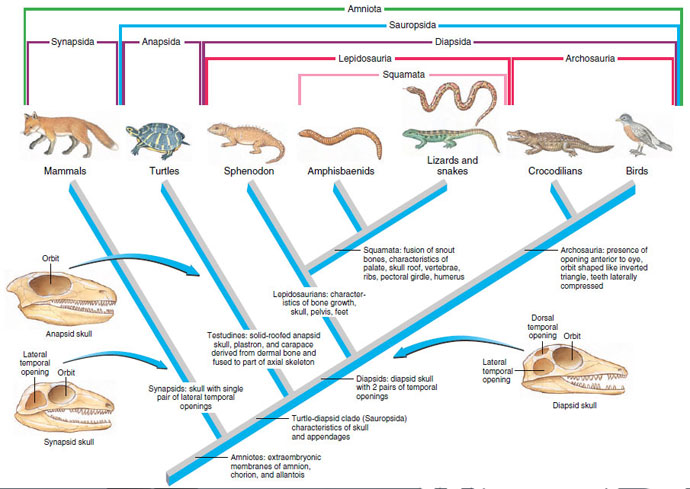 |
| Figure 28-2 Cladogram of the living Amniota showing monophyletic groups. Some of the shared derived characters (synapomorphies) that are diagnostic for the lineages are given. The skulls represent the ancestral condition of the three groups. The skulls of modern diapsids and synapsids are often highly modified by loss or fusion of skull bones that obscures the ancestral condition. Representative skulls for anapsids are Nyctiphruetus of the upper Permian; for diapsids, Youngina of the upper Permian; for synapsids, Aerosaurus, a pelycosaur of the lower Permian. The relationships expressed in this cladogram are tentative and controversial, especially that between birds and mammals. Contrary to the view shown here, in which mammals are the out group, some authorities support a sister-group relationship between birds and mammals based on several kinds of molecular and physiological evidences. |
Characteristics of Class Reptilia
- Body varied in shape, compact in some, elongated in others; body covered with horny epidermal scales with the addition sometimes of bony dermal plates; integument with few glands
- Limbs paired, usually with five toes, and adapted for climbing, running, or paddling; absent in snakes and some lizards
- Skeleton well ossified; ribs with sternum (sternum absent in snakes) forming a complete thoracic basket; skull with one occipital condyle
- Respiration by lungs; no gills; cloaca used for respiration by some; branchial arches in embryonic life
- Three-chambered heart; crocodilians with four-chambered heart; usually one pair of aortic arches; systemic and pulmonary circuits functionally separated
- Ectothermic; many thermoregulate
- Metanephric kidney (paired); uric acid main nitrogenous waste
- Nervous system with the optic lobes on the dorsal side of brain; 12 pairs of cranial nerves in addition to nervus terminalis
- Sexes separate; fertilization internal
- Eggs covered with calcareous or leathery shells; extraembryonic membranes (amnion, chorion, and allantois) present during embryonic life; no aquatic larval stages
Support our developers




