Phylogeny and Adaptive Radiation
Phylogeny and
Adaptive Radiation
Phylogeny
There can be little doubt that the bilaterally symmetrical animals were derived from a radial ancestor, perhaps one very similar to the planula larva of cnidarians. Some investigators believe that this planuloid ancestor may have given rise to one branch of descendants that were sessile or free floating and radial, which became the Cnidaria, and another branch that acquired a creeping habit and bilateral symmetry. Bilateral symmetry is a selective advantage for creeping or swimming animals because sensory structures are concentrated on the anterior end (cephalization), which is the end that first encounters environmental stimuli.
A recent report* cites ribosomal DNA sequences, embryonic cleavage patterns and mesodermal origins, and nervous system structure as evidence that acoels are not, in fact, members of phylum Platyhelminthes. According to the sequence data, acoels are the sister group to all other Bilateria. If this arrangement is sound, then Platyhelminthes as currently constituted is polyphyletic. Relationships of Nemertea and Gnathostomulida are not clear, but they apparently belong in the Lophotrochozoa, along with the remaining flatworms and numerous other phyla.
It seems clear that Turbellaria, even excluding Acoela, is paraphyletic, but we are retaining the taxon for the present because presentation based on thorough cladistic analysis would require introduction of many more taxa and characteristics beyond the scope of this sections. For example, ectolecithal turbellarians should be allied with trematodes, monogeneans, and cestodes in a sister group to endolecithal turbellarians. Some ectolecithal turbellarians share a number of other derived characters with trematodes and cestodes and have been placed by Brooks (1989) in a group designated Cercomeria (Gr. kerkos, tail, + meros, part) (Figure 14-29). Several synapomorphies, including the unique architecture of the tegument, indicates that neodermatans (trematodes, monogeneans, and cestodes) form a monophyletic group.
Adaptive Radiation
The flatworm body plan, with its creeping adaptation, placed a selective advantage on bilateral symmetry and further development of cephalization, ventral and dorsal regions, and caudal differentiation. Because of their body shape and metabolic requirements, early flatworms must have been predisposed toward parasitism and gave rise to symbiotic descendants in the Neodermata. These descendants radiated abundantly as parasites, and many flatworms became very highly specialized for that mode of existence.
Ribbon worms have stressed the proboscis apparatus in their evolutionary diversity. Its use in capturing prey may have been secondarily evolved from its original function as a highly sensitive organ for exploring the environment. Although ribbon worms have evolved beyond flatworms in their complexity of organization, they have been dramatically less abundant as a group. Perhaps the proboscis was so efficient as a predator tool that there was little selective pressure to explore parasitism, or perhaps some critical preconditions were simply not present.
Likewise, jaw worms have not radiated nor become nearly as abundant or diverse as flatworms. However, they have exploited the marine interstitial environment, particularly zones of very low oxygen concentration.
Phylogeny
There can be little doubt that the bilaterally symmetrical animals were derived from a radial ancestor, perhaps one very similar to the planula larva of cnidarians. Some investigators believe that this planuloid ancestor may have given rise to one branch of descendants that were sessile or free floating and radial, which became the Cnidaria, and another branch that acquired a creeping habit and bilateral symmetry. Bilateral symmetry is a selective advantage for creeping or swimming animals because sensory structures are concentrated on the anterior end (cephalization), which is the end that first encounters environmental stimuli.
A recent report* cites ribosomal DNA sequences, embryonic cleavage patterns and mesodermal origins, and nervous system structure as evidence that acoels are not, in fact, members of phylum Platyhelminthes. According to the sequence data, acoels are the sister group to all other Bilateria. If this arrangement is sound, then Platyhelminthes as currently constituted is polyphyletic. Relationships of Nemertea and Gnathostomulida are not clear, but they apparently belong in the Lophotrochozoa, along with the remaining flatworms and numerous other phyla.
It seems clear that Turbellaria, even excluding Acoela, is paraphyletic, but we are retaining the taxon for the present because presentation based on thorough cladistic analysis would require introduction of many more taxa and characteristics beyond the scope of this sections. For example, ectolecithal turbellarians should be allied with trematodes, monogeneans, and cestodes in a sister group to endolecithal turbellarians. Some ectolecithal turbellarians share a number of other derived characters with trematodes and cestodes and have been placed by Brooks (1989) in a group designated Cercomeria (Gr. kerkos, tail, + meros, part) (Figure 14-29). Several synapomorphies, including the unique architecture of the tegument, indicates that neodermatans (trematodes, monogeneans, and cestodes) form a monophyletic group.
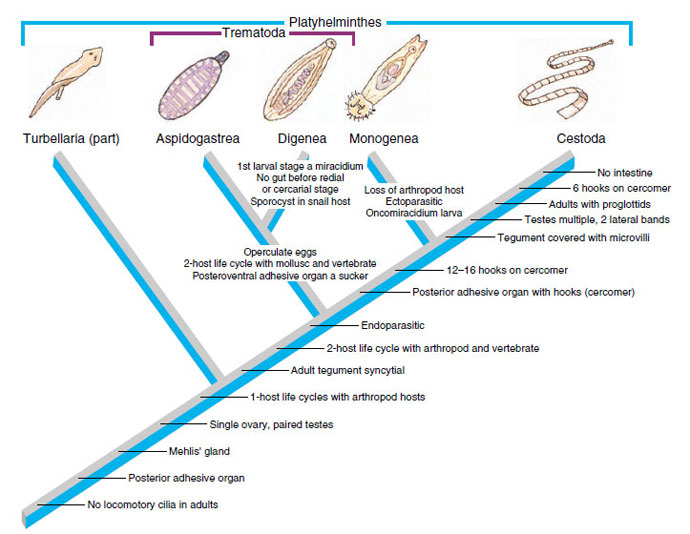 |
| Figure 14-29 Hypothetical relationships among parasitic Platyhelminthes. The traditionally accepted class Turbellaria is paraphyletic. Some turbellarians have ectolecithal development and, together with the Trematoda, Monogenea, and Cestoda, form a clade and a sister group of the endolecithal turbellarians. For the sake of simplicity, the synapomorphies of those turbellarians and of the Aspidogastrea, as well as many others given by Brooks (1989) are omitted. All of these organisms comprise a clade (called Cercomeria) with a posterior adhesive organ. |
Adaptive Radiation
The flatworm body plan, with its creeping adaptation, placed a selective advantage on bilateral symmetry and further development of cephalization, ventral and dorsal regions, and caudal differentiation. Because of their body shape and metabolic requirements, early flatworms must have been predisposed toward parasitism and gave rise to symbiotic descendants in the Neodermata. These descendants radiated abundantly as parasites, and many flatworms became very highly specialized for that mode of existence.
Ribbon worms have stressed the proboscis apparatus in their evolutionary diversity. Its use in capturing prey may have been secondarily evolved from its original function as a highly sensitive organ for exploring the environment. Although ribbon worms have evolved beyond flatworms in their complexity of organization, they have been dramatically less abundant as a group. Perhaps the proboscis was so efficient as a predator tool that there was little selective pressure to explore parasitism, or perhaps some critical preconditions were simply not present.
Likewise, jaw worms have not radiated nor become nearly as abundant or diverse as flatworms. However, they have exploited the marine interstitial environment, particularly zones of very low oxygen concentration.
Support our developers




