Phylum Hemichordata
Phylum
Hemichordata
Hemichordata (hem´i-kor-da´ta) (Gr. hemi, half, + chorda, string, cord) are marine animals that were formerly considered a subphylum of chordates, based on their possession of gill slits and a rudimentary notochord. However, the so-called hemichordate notochord is really a buccal diverticulum (called a stomochord, meaning “mouth-cord”) and not homologous with the chordate notochord, so hemichordates are given the rank of a separate phylum.
Hemichordates are vermiform bottom dwellers, living usually in shallow waters. Some colonial species live in secreted tubes. Most are sedentary or sessile. Their distribution is almost cosmopolitan, but their secretive habits and fragile bodies make collecting them difficult.
Members of class Enteropneusta (Gr. enteron, intestine, + pneustikos, of, or for, breathing) (acorn worms) range from 20 mm to 2.5 m in length. Members of class Pterobranchia (Gr. pteron, wing, + branchia, gills) are smaller, usually 1 to 12 mm, not including the stalk. About 70 species of enteropneusts and two small genera of pterobranchs are recognized.
Hemichordates have the typical tricoelomate structure of deuterostomes.
Class Enteropneusta
Enteropneusts, or acorn worms, are sluggish, wormlike animals that live in burrows or under stones, usually in mud or sand flats of intertidal zones. Balanoglossus (Gr. balanos, acorn, + gl|ssa, tongue) and Saccoglossus (Gr. sakkos, sac, strainer, + glossa, tongue) (Figure 24-2) are common genera.
Form and Function
The mucus-covered body is divided into a tonguelike proboscis, a short collar, and a long trunk (protosome, mesosome, and metasome).
Proboscis: The proboscis is the active part of the animal. It probes about in the mud, examining its surroundings and collecting food in mucous strands on its surface. Cilia carry particles to the groove at the edge of the collar, direct them to the mouth on the underside, and then the particles are swallowed. Large particles can be rejected by covering the mouth with the edge of the collar (Figure 24-3).
Burrow dwellers use the proboscis to excavate, thrusting it into the mud or sand and allowing cilia and mucus to move the sand backward. Or they may ingest sand or mud as they go, extracting its organic contents. They build U-shaped, mucus-lined burrows, usually with two openings 10 to 30 cm apart and with the base of the U 50 to 75 cm below the surface. They can thrust their proboscis out the front opening for feeding. Defecation at the back opening builds characteristic spiral mounds of feces that leave a telltale clue to the location of burrows.
In the posterior end of the proboscis is a small coelomic sac (protocoel) into which extends the buccal diverticulum, a slender, blindly ending pouch of the gut that reaches forward into the buccal region and was formerly considered a notochord. A slender canal connects the protocoel with a proboscis pore to the outside (Figure 24-2B). The paired coelomic cavities in the collar also open by pores. By taking in water through the pores into the coelomic sacs, the proboscis and collar can be stiffened to aid in burrowing. Contraction of the body musculature then forces the excess water out through the gill slits, reducing the hydrostatic pressure and allowing the animal to move forward.
Branchial System: A row of gill pores is located dorsolaterally on each side of the trunk just behind the collar (Figure 24-3A). Pores open from a series of gill chambers that in turn connect with a series of gill slits in the sides of the pharynx. There are no gills on the gill slits, but some respiratory gaseous exchange occurs in the vascular branchial epithelium, as well as in the body surface. Ciliary currents keep a fresh supply of water moving from the mouth through the pharynx and out the gill slits and branchial chambers to the outside.
Feeding and the Digestive System: Hemichordates are largely ciliary-mucus feeders. Behind the buccal cavity lies the large pharynx containing in its dorsal part the U-shaped gill slits (Figure 24-2B). Since there are no gills, the primary function of the branchial mechanism of the pharynx is presumably food gathering. Having been caught in mucus and brought to the mouth by ciliary action on the proboscis and collar, food particles are strained from the branchial water that leaves through the gill slits. Food then passes to the ventral part of the pharynx and esophagus to the intestine, where digestion and absorption occur (Figure 24-3).
Circulatory and Excretory Systems A middorsal vessel carries the colorless blood forward above the gut. In the collar the vessel expands into a sinus and a heart vesicle above the buccal diverticulum. Blood then enters a network of blood sinuses called the glomerulus, which partially surrounds these structures. The glomerulus is assumed to have an excretory function (Figure 24-2B). Blood travels posteriorly through a ventral vessel below the gut, passing through extensive sinuses to the gut and body wall.
Nervous and Sensory Systems: The nervous system consists mostly of a subepithelial network, or plexus, of nerve cells and fibers to which processes of epithelial cells are attached. Thickenings of this net form dorsal and ventral nerve cords that are united posterior to the collar by a ring connective. The dorsal cord continues into the collar and furnishes many fibers to the plexus of the proboscis. The collar cord is hollow in some species and contains giant nerve cells with processes running to the nerve trunks. This nerve plexus system is quite reminiscent of that of cnidarians and echinoderms.
Sensory receptors include neurosensory cells throughout the epidermis (especially in the proboscis, a preoral ciliary organ that may be chemoreceptive) and photoreceptor cells.
Reproductive System and Development: Sexes are separate in enteropneusts. A dorsolateral row of gonads runs along each side of the anterior part of the trunk. Fertilization is external, and in some species a ciliated tornaria larva develops that at certain stages is so similar to the echinoderm bipinnaria that it was once believed to be an echinoderm larva (Figure 24-4). The familiar Saccoglossus of American waters has direct development without a tornaria stage.
Characteristics of Phylum Hemichordata
Class Pterobranchia
The basic plan of class Pterobranchia is similar to that of Enteropneusta, but certain structural differences are correlated with the sedentary life-style of pterobranchs. The first pterobranch ever reported was obtained by the famed Challenger expedition of 1872 to 1876. Although first placed among Polyzoa (Entoprocta and Ectoprocta), its affinities to hemichordates were later recognized. Only two genera (Cephalodiscus and Rhabdopleura) are known in any detail.
Pterobranchs are small animals, usually within the range of 1 to 7 mm in length, although the stalk may be longer. Many individuals of Cephalodiscus (Gr. kephale, head, + diskos, disc) (Figure 24-5) live together in collagenous tubes, which often form an anastomosing system. Zooids are not connected, however, and live independently in the tubes. Through apertures in these tubes, they extend their crown of tentacles. They are attached to the walls of the tubes by extensible stalks that can jerk the owners back into the tubes when necessary.
The body of Cephalodiscus is divided into the three regions—proboscis, collar, and trunk—characteristic of hemichordates. There is only one pair of gill slits, and the alimentary canal is U-shaped, with the anus near the mouth. The proboscis is shield shaped. At the base of the proboscis are five to nine pairs of branching arms with tentacles containing an extension of the coelomic compartment of the mesosome, as in a lophophore. Ciliated grooves on the tentacles and arms collect food. Some species are dioecious, and others are monoecious. Asexual reproduction by budding may also occur.
In Rhabdopleura (Gr. rhabdos, rod, + pleura, a rib, the side), which is smaller than Cephalodiscus, the members remain together to form a colony of zooids connected by a stolon and enclosed in secreted tubes (Figure 24-6). The collar in these forms bears two branching arms. No gill clefts or glomeruli are present. New individuals are produced by budding from a creeping basal stolon, which branches on a substratum. No pterobranch has a tubular nerve cord in the collar, but otherwise their nervous system is similar to that of Enteropneusta.
The fossil graptolites of the middle Paleozoic era often are placed as an extinct class under Hemichordata. They are important index fossils of the Ordovician and Silurian geological strata. Alignment of graptolites with the hemichordates has been very controversial, but discovery of an organism that seems to be a living graptolite lends strong support to the hypothesis. It has been described as a new species of pterobranch, called Cephalodiscus graptolitoides.
Hemichordata (hem´i-kor-da´ta) (Gr. hemi, half, + chorda, string, cord) are marine animals that were formerly considered a subphylum of chordates, based on their possession of gill slits and a rudimentary notochord. However, the so-called hemichordate notochord is really a buccal diverticulum (called a stomochord, meaning “mouth-cord”) and not homologous with the chordate notochord, so hemichordates are given the rank of a separate phylum.
Hemichordates are vermiform bottom dwellers, living usually in shallow waters. Some colonial species live in secreted tubes. Most are sedentary or sessile. Their distribution is almost cosmopolitan, but their secretive habits and fragile bodies make collecting them difficult.
Members of class Enteropneusta (Gr. enteron, intestine, + pneustikos, of, or for, breathing) (acorn worms) range from 20 mm to 2.5 m in length. Members of class Pterobranchia (Gr. pteron, wing, + branchia, gills) are smaller, usually 1 to 12 mm, not including the stalk. About 70 species of enteropneusts and two small genera of pterobranchs are recognized.
Hemichordates have the typical tricoelomate structure of deuterostomes.
Class Enteropneusta
Enteropneusts, or acorn worms, are sluggish, wormlike animals that live in burrows or under stones, usually in mud or sand flats of intertidal zones. Balanoglossus (Gr. balanos, acorn, + gl|ssa, tongue) and Saccoglossus (Gr. sakkos, sac, strainer, + glossa, tongue) (Figure 24-2) are common genera.
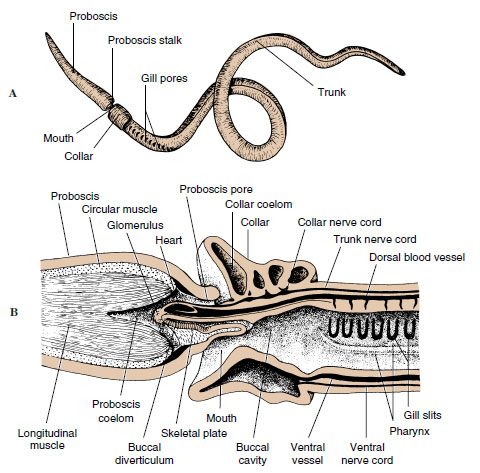 |
| Figure 24-2 Acorn worm Saccoglossus (Hemichordata, class Enteropneusta). A, External lateral view. B, Longitudinal section through anterior end. |
Form and Function
The mucus-covered body is divided into a tonguelike proboscis, a short collar, and a long trunk (protosome, mesosome, and metasome).
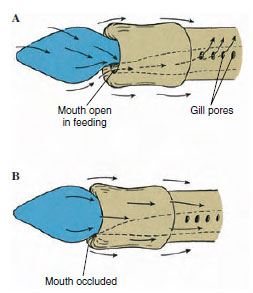 |
| Figure 24-3 Food currents of enteropneust hemichordate. A, Side view of acorn worm with mouth open, showing direction of currents created by cilia on proboscis and collar. Food particles are directed toward mouth and digestive tract. Rejected particles move toward outside of collar. Water leaves through gill pores. B, When mouth is occluded, all particles are rejected and passed onto the collar. Nonburrowing and some burrowing hemichordates use this feeding method. |
Proboscis: The proboscis is the active part of the animal. It probes about in the mud, examining its surroundings and collecting food in mucous strands on its surface. Cilia carry particles to the groove at the edge of the collar, direct them to the mouth on the underside, and then the particles are swallowed. Large particles can be rejected by covering the mouth with the edge of the collar (Figure 24-3).
Burrow dwellers use the proboscis to excavate, thrusting it into the mud or sand and allowing cilia and mucus to move the sand backward. Or they may ingest sand or mud as they go, extracting its organic contents. They build U-shaped, mucus-lined burrows, usually with two openings 10 to 30 cm apart and with the base of the U 50 to 75 cm below the surface. They can thrust their proboscis out the front opening for feeding. Defecation at the back opening builds characteristic spiral mounds of feces that leave a telltale clue to the location of burrows.
In the posterior end of the proboscis is a small coelomic sac (protocoel) into which extends the buccal diverticulum, a slender, blindly ending pouch of the gut that reaches forward into the buccal region and was formerly considered a notochord. A slender canal connects the protocoel with a proboscis pore to the outside (Figure 24-2B). The paired coelomic cavities in the collar also open by pores. By taking in water through the pores into the coelomic sacs, the proboscis and collar can be stiffened to aid in burrowing. Contraction of the body musculature then forces the excess water out through the gill slits, reducing the hydrostatic pressure and allowing the animal to move forward.
Branchial System: A row of gill pores is located dorsolaterally on each side of the trunk just behind the collar (Figure 24-3A). Pores open from a series of gill chambers that in turn connect with a series of gill slits in the sides of the pharynx. There are no gills on the gill slits, but some respiratory gaseous exchange occurs in the vascular branchial epithelium, as well as in the body surface. Ciliary currents keep a fresh supply of water moving from the mouth through the pharynx and out the gill slits and branchial chambers to the outside.
Feeding and the Digestive System: Hemichordates are largely ciliary-mucus feeders. Behind the buccal cavity lies the large pharynx containing in its dorsal part the U-shaped gill slits (Figure 24-2B). Since there are no gills, the primary function of the branchial mechanism of the pharynx is presumably food gathering. Having been caught in mucus and brought to the mouth by ciliary action on the proboscis and collar, food particles are strained from the branchial water that leaves through the gill slits. Food then passes to the ventral part of the pharynx and esophagus to the intestine, where digestion and absorption occur (Figure 24-3).
Circulatory and Excretory Systems A middorsal vessel carries the colorless blood forward above the gut. In the collar the vessel expands into a sinus and a heart vesicle above the buccal diverticulum. Blood then enters a network of blood sinuses called the glomerulus, which partially surrounds these structures. The glomerulus is assumed to have an excretory function (Figure 24-2B). Blood travels posteriorly through a ventral vessel below the gut, passing through extensive sinuses to the gut and body wall.
Nervous and Sensory Systems: The nervous system consists mostly of a subepithelial network, or plexus, of nerve cells and fibers to which processes of epithelial cells are attached. Thickenings of this net form dorsal and ventral nerve cords that are united posterior to the collar by a ring connective. The dorsal cord continues into the collar and furnishes many fibers to the plexus of the proboscis. The collar cord is hollow in some species and contains giant nerve cells with processes running to the nerve trunks. This nerve plexus system is quite reminiscent of that of cnidarians and echinoderms.
Sensory receptors include neurosensory cells throughout the epidermis (especially in the proboscis, a preoral ciliary organ that may be chemoreceptive) and photoreceptor cells.
Reproductive System and Development: Sexes are separate in enteropneusts. A dorsolateral row of gonads runs along each side of the anterior part of the trunk. Fertilization is external, and in some species a ciliated tornaria larva develops that at certain stages is so similar to the echinoderm bipinnaria that it was once believed to be an echinoderm larva (Figure 24-4). The familiar Saccoglossus of American waters has direct development without a tornaria stage.
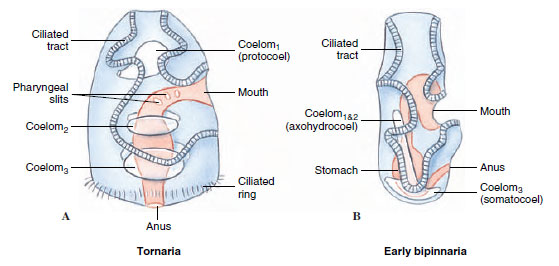 |
| Figure 24-4 Comparison of a hemichordate tornaria (A) to an echinoderm bipinnaria (B). |
Characteristics of Phylum Hemichordata
- Soft bodied; wormlike or short and compact with stalk for attachment
- Body divided into proboscis, collar, and trunk; coelomic pouch single in proboscis, but paired in other two; buccal diverticulum in posterior part of proboscis
- Enteropneusta free moving and of burrowing habits; pterobranchs sessile, mostly colonial, living in secreted tubes
- Circulatory system of dorsal and ventral vessels and dorsal heart
- Respiratory system of gill slits (few or none in pterobranchs) connecting the pharynx with outside as in chordates
- No nephridia; a single glomerulus connected to blood vessels may have excretory function
- A subepidermal nerve plexus thickened to form dorsal and ventral nerve cords, with a ring connective in the collar; dorsal nerve cord of collar hollow in some
- Sexes separate in Enteropneusta, with gonads projecting into body cavity; in pterobranchs reproduction may be sexual or asexual (in some) by budding; tornaria larva in some Enteropneusta
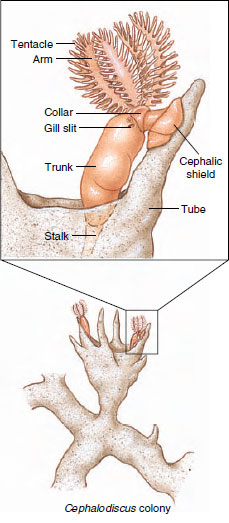 |
| Figure 24-5 Cephalodiscus, a pterobranch hemichordate. These tiny (5 to 7 mm) forms live in tubes in which they can move freely. Ciliated tentacles and arms direct currents of food and water toward mouth |
Class Pterobranchia
The basic plan of class Pterobranchia is similar to that of Enteropneusta, but certain structural differences are correlated with the sedentary life-style of pterobranchs. The first pterobranch ever reported was obtained by the famed Challenger expedition of 1872 to 1876. Although first placed among Polyzoa (Entoprocta and Ectoprocta), its affinities to hemichordates were later recognized. Only two genera (Cephalodiscus and Rhabdopleura) are known in any detail.
Pterobranchs are small animals, usually within the range of 1 to 7 mm in length, although the stalk may be longer. Many individuals of Cephalodiscus (Gr. kephale, head, + diskos, disc) (Figure 24-5) live together in collagenous tubes, which often form an anastomosing system. Zooids are not connected, however, and live independently in the tubes. Through apertures in these tubes, they extend their crown of tentacles. They are attached to the walls of the tubes by extensible stalks that can jerk the owners back into the tubes when necessary.
The body of Cephalodiscus is divided into the three regions—proboscis, collar, and trunk—characteristic of hemichordates. There is only one pair of gill slits, and the alimentary canal is U-shaped, with the anus near the mouth. The proboscis is shield shaped. At the base of the proboscis are five to nine pairs of branching arms with tentacles containing an extension of the coelomic compartment of the mesosome, as in a lophophore. Ciliated grooves on the tentacles and arms collect food. Some species are dioecious, and others are monoecious. Asexual reproduction by budding may also occur.
In Rhabdopleura (Gr. rhabdos, rod, + pleura, a rib, the side), which is smaller than Cephalodiscus, the members remain together to form a colony of zooids connected by a stolon and enclosed in secreted tubes (Figure 24-6). The collar in these forms bears two branching arms. No gill clefts or glomeruli are present. New individuals are produced by budding from a creeping basal stolon, which branches on a substratum. No pterobranch has a tubular nerve cord in the collar, but otherwise their nervous system is similar to that of Enteropneusta.
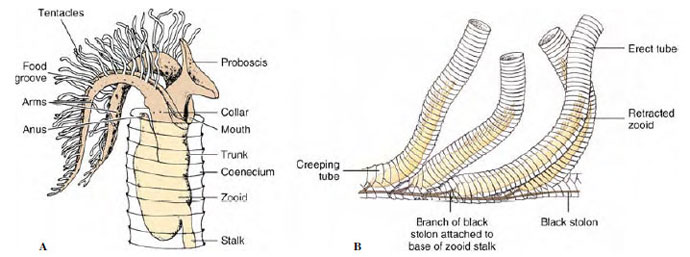 |
| Figure 24-6 A, Rhabdopleura, a pterobranch hemichordate in its tube. Individuals live in branching tubes connected by stolons, and protrude the ciliated tentacles for feeding. B, Portion of a colony. |
The fossil graptolites of the middle Paleozoic era often are placed as an extinct class under Hemichordata. They are important index fossils of the Ordovician and Silurian geological strata. Alignment of graptolites with the hemichordates has been very controversial, but discovery of an organism that seems to be a living graptolite lends strong support to the hypothesis. It has been described as a new species of pterobranch, called Cephalodiscus graptolitoides.
Support our developers




